Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Заключение
Теперь мы можем подвести итоги. С появлением белкового синтеза первые живые системы сильно расширили свои возможности обмена веществ. Первая белковая РНК-зависимая РНК-полимераза сняла с рибозимов необходимость заниматься собственной репликацией и позволила сильно увеличить количество хранимой генетической информации. Белки также стали промежуточным звеном, создавшим возможность строить липидные мембраны. Так жизнь перешла из плоской формы прилипших к минеральной поверхности РНК к трехмерной форме – скоплениям РНК и белков во впадинах и полостях минеральных отложений, закрытых примитивными мембранами. Полная независимость от глины, сульфида цинка и других минералов тогда была еще невозможна, но появились первые структуры, похожие на клетки, – закрытые мембраной со всех сторон пузырьки, плавающие в воде. Они поначалу были расселительными стадиями плоских, сидящих на минералах протоорганизмов, геном которых состоял, по всей видимости, из множества разных молекул РНК, одно– или двухцепочечных, линейных и кольцевых. Механизмы их репликации, скорее всего, напоминали таковые у РНК-вирусов, их размер соответствовал размеру геномов РНК-вирусов, в пределах 30 000 нуклеотидов. Геномная молекула такого размера может кодировать простую систему трансляции с ее рРНК, тРНК, рибосомными белками и другими факторами.
Возможно, в протоорганизмах существовала «доминирующая» молекула РНК, кодирующая РНК-полимеразу и главные компоненты рибосомы, и многочисленные РНК-плазмиды, кодирующие ферменты обмена веществ, средства защиты и взаимодействия с внешним миром. При делении протоорганизмов эти геномные молекулы расходились не поровну и не попадали в некоторых потомков, что компенсировалось их способностью выходить наружу в виде вирусоподобных частиц и проникать в другие генетические комплексы. Наряду с «полезными» плазмидами уже тогда должен был быть весь спектр эгоистичных генетических элементов: от маленьких, ничего не кодирующих вироидов и специализированных вирусов с белковыми оболочками до внутригеномных паразитов, таких как интроны I типа и частично эгоистичных плазмид.
Вирусы в ходе «гонки вооружений» со своими хозяевами – плоскими протоорганизмами создали новые типы нуклеиновых кислот, такие как метил-РНК, урацил-ДНК и современную ДНК с тимином. Эти новые нуклеиновые кислоты были затем подхвачены протоорганизмами и позволили увеличить размер и стабильность генома. Изобретение ДНК и совершенствование механизмов ее копирования происходило параллельно в разных линиях вирусов, что привело к большому разнообразию ферментов, работающих с ДНК. Наконец, появление надежной репликации ДНК на матрице ДНК подготовило почву для объединения разнообразных генетических элементов в единые большие ДНК-геномы и последующего исхода бактериальных и архейных клеток из геотермальной колыбели в большой мир.
Глава 15
Происхождение мембран и мембранной энергетики
Какие мембраны были у LUCA?
В прошлой главе мы проследили происхождение геномной ДНК и механизмов ее репликации – от РНК-белкового мира до больших кольцевых молекул ДНК, составляющих геномы бактерий и архей. Попробуем теперь путем сравнения бактерий и архей узнать структуру мембран их общего предка.
Все клетки окружены мембраной – тонкой подвижной пленкой из двух слоев особых молекул, которые называются липидами. Почему липиды образуют именно пленку, а не капли? Схожие механизмы формируют стенки мыльных пузырей и масляные пленки на поверхности воды.
Чтобы разобраться в этом, надо обратиться к деталям строения молекул разных веществ. Например, в молекуле воды электроны, образующие связи кислорода с водородом, смещены к атому кислорода. Поэтому молекула воды электрически полярна – одна ее сторона несет положительный электрический заряд, а другая – отрицательный. Благодаря полярности молекулы воды притягиваются друг к другу. Другие вещества с полярными молекулами, такие как аммиак и этиловый спирт, прекрасно смешиваются с водой.
Если молекула состоит из атомов с примерно одинаковой электроотрицательностью, то в ней не будет местных электрических зарядов. Такими свойствами обладают, например, углеводороды (метан СН 4, октан С 8Н 18, бензол С 6Н 6и др.). Неполярные молекулы не притягиваются к полярным. Поэтому при смешивании полярной жидкости с неполярной, например масла с водой, получается не раствор, а эмульсия – взвесь капель одной жидкости в другой. Если в воде или в масле перед смешиванием были растворены какие-либо вещества, они могут переходить из одной жидкости в другую, более подходящую ей по полярности молекул. Это хорошо видно в тарелке борща: красный краситель свеклы (беталаин) имеет полярные молекулы и растворяется в воде, а желтый краситель моркови (каротин) неполярен и переходит в капли жира на поверхности (рис. 15.1). Полярные молекулы еще называются гидрофильными («любящими воду»), а неполярные – гидрофобными («боящимися воды»).
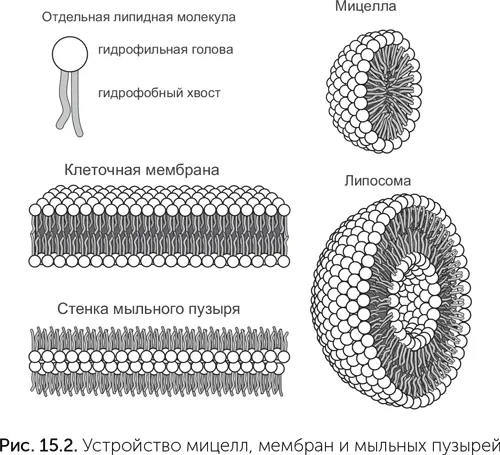
Для получения мембран и мыльных пузырей нужны молекулы с более сложными свойствами. Они должны быть вытянутой формы, с одним полярным концом и другим неполярным. Простейшие молекулы с такими свойствами – жирные кислоты. Их натриевые соли широко используются под названием «мыло». При растворении мыла в воде его молекулы образуют мельчайшие, нанометрового размера шарики и палочки – мицеллы (рис. 15.2). Каждая молекула в мицелле полярным концом контактирует с водой, а неполярный конец спрятан внутри. На поверхности воды мыло образует слой толщиной в одну молекулу. Полярные концы молекул мыла обращены в воду, а неполярные – к воздуху. Стенка мыльного пузыря состоит из двух слоев молекул, они собраны полярными концами внутрь, а неполярными – к воздуху по обе стороны от стенки. Наконец, клеточная мембрана похожа на стенку мыльного пузыря, только вывернутую наизнанку. В мембране полярные концы липидных молекул обращены к воде по обе стороны, а неполярные скрыты внутри.
Мембраны современных бактерий состоят из фосфолипидов – сложных эфиров глицерола, двух остатков жирной кислоты и одного фосфатного остатка (рис. 15.3). К фосфатному остатку может быть присоединена дополнительная полярная группа: это может быть этаноламин, холин, аминокислота серин или многоатомный спирт инозитол. Гидрофобные хвосты жирных кислот образуют средний слой мембраны, а полярные остатки глицерола, фосфата и вспомогательных полярных групп – наружный и внутренний слои. Мембраны архей устроены в принципе похоже, но на другой химической основе. Их липиды имеют в качестве гидрофобной части терпеновые спирты, например геранилгераниол [13]. Углеводородные цепочки терпенов несут метильные (СН 3) группы через каждые четыре атома. Эти спирты простыми эфирными связями присоединяются к глицерол-фосфату, к его фосфатному остатку тоже могут присоединяться дополнительные полярные головки, такие же, как у бактерий. Глицерол-фосфат архей тоже отличается от бактериального – у архей используется другой его оптический изомер, глицерол-1-фосфат вместо глицерол-3-фосфата.
Читать дальшеИнтервал:
Закладка:






