Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Мембранная электрохимия
Важнейшей функцией мембран является поддержание мембранного электрического потенциала, который используется клеткой для запасания и передачи энергии. Клеточная мембрана работает как конденсатор: ее наружная сторона несет положительный заряд, а внутренняя – отрицательный. Разность потенциалов между сторонами мембраны в активном состоянии клетки может достигать 150–200 милливольт. Напряжение, казалось бы, небольшое, но оно приложено к слою изолятора толщиной всего 10 нанометров, т. е. клеточная мембрана – хороший изолятор. Заряда этого конденсатора хватает на несколько секунд активной жизнедеятельности клетки.
Мембранные электрохимические процессы используются для синтеза основной части клеточного АТФ у большинства современных организмов. В большинстве случаев, в том числе в клетках человека, энергия для получения АТФ в конечном итоге получается в реакциях окисления сахаров и жиров кислородом, т. е. в клеточном дыхании. На первых этапах дыхания (в случае сахаров это гликолиз и цикл Кребса, мы обсуждали их в 11–1 главе) сахара расщепляются, все атомы углерода из их молекул переходят в углекислый газ, а все атомы водорода – на носители, главным образом НАД:
С 6Н12О 6(глюкоза) + 6 Н 2О + 10 НАД ++ 2 ФАД → 6 СО 2 + 10 НАДН + 10 Н ++ 2 ФАДН 2.
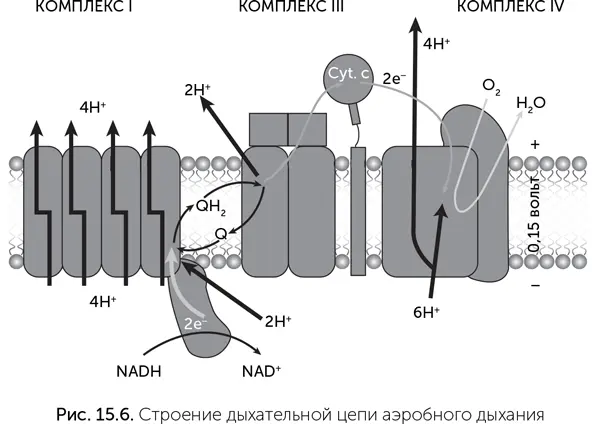
Эта часть дыхания происходит без участия мембран и кислорода. Мембраны вступают в игру на последующих этапах, а кислород – только в самом конце. Основная часть АТФ образуется благодаря работе белков так называемой «дыхательной цепи», которые находятся в мембране (рис. 15.6). Дыхательная цепь состоит из больших белковых комплексов (они называются неоригинально: комплекс I, комплекс II, III и IV) и маленьких подвижных переносчиков электронов (кофермент Q и цитохром с ). Эти молекулы по цепочке передают электроны от НАДН и ФАДН 2на кислород. Основной поток электронов идет следующим путем: НАДН → комплекс I → кофермент Q → комплекс III → цитохром с → комплекс IV → кислород. Комплекс II играет вспомогательную роль, через него электроны с ФАДН 2входят в дыхательную цепь на кофермент Q.
При прохождении электронов через комплексы дыхательной цепи выделяется энергия, которая используется на откачку из клетки наружу ионов водорода – протонов (Н +). Далее протоны входят обратно в клетку через другой белок – мембранную АТФазу, которая за счет энергии протонов делает АТФ из АДФ и фосфорной кислоты.
Дыхательная цепь называется еще «электрон-транспортной цепью», потому что в ней происходит передача электронов от одних молекул к другим по цепочке.
Напряжение на мембране используется не только для синтеза АТФ, но и для транспорта нужных веществ в клетку. Поскольку на наружной стороне мембраны образуется положительный заряд, то для транспорта незаряженных молекул, таких как сахара, нужно организовать их движение в клетку совместно с протонами. Транспортный белок должен связать на наружной стороне мембраны молекулу сахара и протон, затем изогнуться или вывернуться так, чтобы сахар и протон оказались на внутренней стороне мембраны, и выпустить их внутрь клетки. Для транспорта молекул с отрицательным зарядом, таких как фосфат и нуклеотиды, вместе с каждой нужной молекулой нужно впускать в клетку несколько протонов, чтобы скомпенсировать отрицательный заряд «полезного груза».
Происхождение мембранных АТФаз
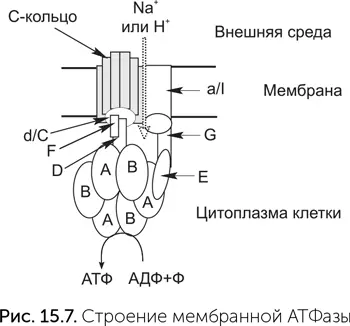
Комплексы дыхательных цепей очень разнообразны, подстраиваются к условиям обитания клетки, и их гены подвержены горизонтальному переносу. Мы обсудим их происхождение в главе 17. Мембранные АТФазы же во всех клетках устроены довольно похоже. Молекулы мембранной АТФазы состоят из вращающейся части («ротор») и неподвижной («статор») (рис. 15.7). «Ротор» состоит из кольца из 8–15 мелких субъединиц С в мембране, центрального стержня из субъединиц D, d/C и F в цитоплазме. В состав статора входят боковой стержень, выступающий в цитоплазму (субъединицы E и G), и кольцо из трех А– и трех В-субъединиц. Центральный стержень ротора входит в кольцо из А и В-субъединиц, и при вращении его конец изгибает и сдвигает субъединицы А и В друг относительно друга.
Подробное видео работы АТФазы можно посмотреть по адресу: http://haa.su/Av9/.
Протоны связываются двумя карманами «статора» – по одному с каждой стороны мембраны, и чтобы пройти мембрану, они должны, связавшись с С-субъединицей, ждать поворота C-кольца на одну субъединицу. Это единственный путь от внутреннего «кармана» статора к наружному. Молекулы ATФ и АДФ связываются между A и В субъединицами цитоплазматического «статора», активный АТФазный центр образуется в контакте со статором. Чтобы из АДФ и фосфата получилась молекула АТФ, нужно движение А-субъединицы относительно соседней В-субъединицы. Это движение обеспечивает ротор за счет энергии проходящих через АТФазу протонов. На один оборот ротора каждый из трех АТФазных центров АВ-кольца создает одну молекулу АТФ. Таким образом, на один оборот молекулы через мембрану переносится от 8 до 15 протонов (по одному на каждую C-субъединицу) и синтезируется три молекулы АТФ.
Важнейшее преимущество такого преобразования энергии состоит в том, что роторная АТФаза позволяет клетке использовать для синтеза АТФ химические реакции, выделяющие мало энергии. Она синтезирует за один оборот три молекулы АТФ, а количество прошедших ионов равно числу С-субъединиц – от 8 до 15. Иначе говоря, на синтез одной молекулы АТФ приходится от трех до пяти прошедших ионов. Роторная АТФаза работает как своего рода повышающий трансформатор: достаточно, чтобы выделяющейся в химической реакции энергии хватило на перенос через мембрану одного протона, дальше из множества этих маленьких порций энергии будет собрано несколько более крупных.
Основные субъединицы АТФазы и их взаимное расположение похожи на систему секреции белков III типа (рис. 15.8). Эта общая для бактерий и архей система активно выделяет белки из клетки, развернув их в цепочку, с затратой АТФ и вращением выделяемого белка. А– и В-субъединицы статора также обладают более далеким, но достоверным сходством с хеликазами семейства Rho и белком RecA. Эти белки образуют кольца из шести одинаковых субъединиц, которые вращаются вокруг ДНК, расплетая ее с затратой АТФ (обсуждалось в главе 14). Таким образом, мембранная АТФаза происходит от древнего семейства вращающихся АТФаз с разнообразными функциями. Вероятно, сначала предковая РНК-хеликаза вступила в контакт с мембранной порой и стала работать системой экспорта РНК из протоклетки. АВ-кольцо, происходящее от хеликазы, не только проталкивало экспортируемую РНК наружу, но и крутило ее. Потом этот белковый комплекс переключился с РНК на белки и дал начало системе секреции белков III типа. Потом секретируемый белок застрял в поре этой системы секреции и стал передавать вращение, создаваемое АВ-кольцом, на С-кольцо. Для возникновения роторной АТФ-синтазы осталось только совместить вращение белка и транспорт ионов через него. В составе С-кольца системы секреции белков III типа уже есть участки связывания ионов натрия, которые помогают скреплению С-субъединиц друг с другом. Мутации бокового стержня статора могли создать проходы для этих ионов к обеим сторонам мембраны и превратить заклинившую систему секреции в роторную АТФазу.
Читать дальшеИнтервал:
Закладка:






