Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
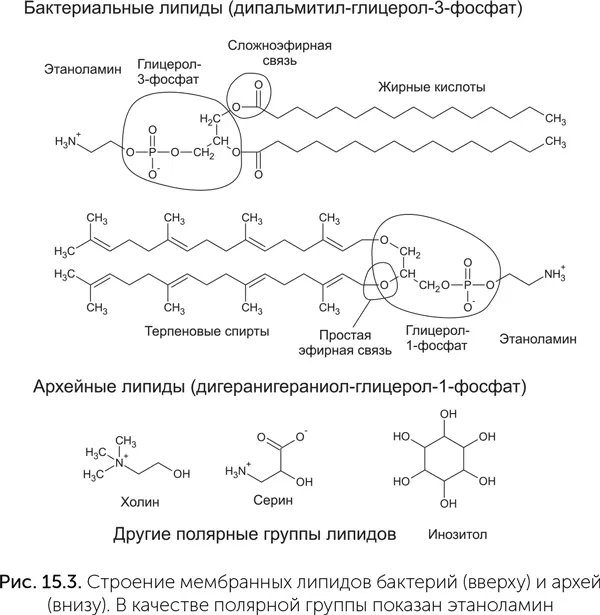
Таким образом, сравнение современных мембран ничего нам не дает для понимания их общего предкового состояния – все основные компоненты отличаются вплоть до полной несовместимости. Одна из крайних точек зрения, высказанная Мартином и Расселом (Martin, Russell, 2007), заключается в том, что последний общий предок не имел мембран вовсе и современные мембраны были изобретены независимо предками бактерий и архей при выходе из минеральных каверн.
Другая крайняя точка зрения основана на обнаружении жирных кислот в метеоритах и в условиях опыта Миллера. Согласно ей примитивные мембраны, состоящие из абиогенно синтезированных жирных кислот, существовали еще на заре мира РНК, до появления белков. Обе эти крайности, скорее всего, неверны. В реконструированном арсенале белков LUCA есть несколько мембранных белков, таких как роторная мембранная АТФаза и система секреции белков III типа. Они не могли бы сформироваться без существования хоть каких-нибудь мембран. Абиогенные жирные кислоты же обладают большим разбросом в длине углеводородной цепи, и поэтому из их смеси получаются крайне непрочные мембраны. Вахтерхойзер предполагал, что LUCA имел смесь липидов с обоими изомерами глицерола, а бактерии и археи унаследовали по одному типу из этих двух (Wächtershäuser, 2006). Однако, когда такие мембраны были получены искусственно, оказалось, что липиды с разными изомерами глицерола быстро разделяются на «острова», содержащие преимущественно один изомер из двух, а мембрана легко рвется по границам этих «островов».
Вопрос происхождения мембран также требует решения очередного парадокса «курицы и яйца»: современные мембраны непроницаемы для ионов металлов и заряженных органических молекул, таких как аминокислоты, и слабо пропускают сахара. Чтобы клетки могли поглощать органические вещества из внешней среды, мембрана содержит десятки видов транспортных белков. Клетка с мембраной без транспортных белков обречена на голод, а транспортные белки не могут возникнуть в отсутствие мембран. Хуже того, синтез мембранных белков в современных клетках требует участия мембранного белкового комплекса SRP, который связывается с рибосомой и помогает встроить в мембрану выходящую из нее белковую цепь. Без SRP участок пептида, который должен быть в мембране и состоит из неполярных аминокислот, просто застревает на выходе из рибосомы (Mulkidjanian, Galperin, Koonin, 2009)!
Информация о мембранах LUCA сохранилась в ферментах синтеза липидов
Если сами липиды бактерий и архей сильно различаются, вплоть до полной неузнаваемости, то некоторые ферменты, участвующие в их синтезе, довольно похожи. Синтез липидов состоит из многих этапов (рис. 15.4). Сначала образуются гидрофобные хвосты. Жирные кислоты собираются из ацетил-КоА, на каждом шаге синтеза молекула жирной кислоты присоединяет один ацетильный (СН 3-СО) фрагмент и вырастает на два атома углерода. Терпены тоже образуются из ацетил-КоА. На первой стадии из трех молекул ацетил-КоА образуется мевалоновая кислота, которая превращается в изопентил-пирофосфат с пятью атомами углерода. Потом из молекул изопентил-пирофосфата собираются более длинные терпены: геранил-пирофосфат, фарнезил-пирофосфат и геранилгеранил-пирофосфат, содержащие 10, 15 и 20 атомов углерода.
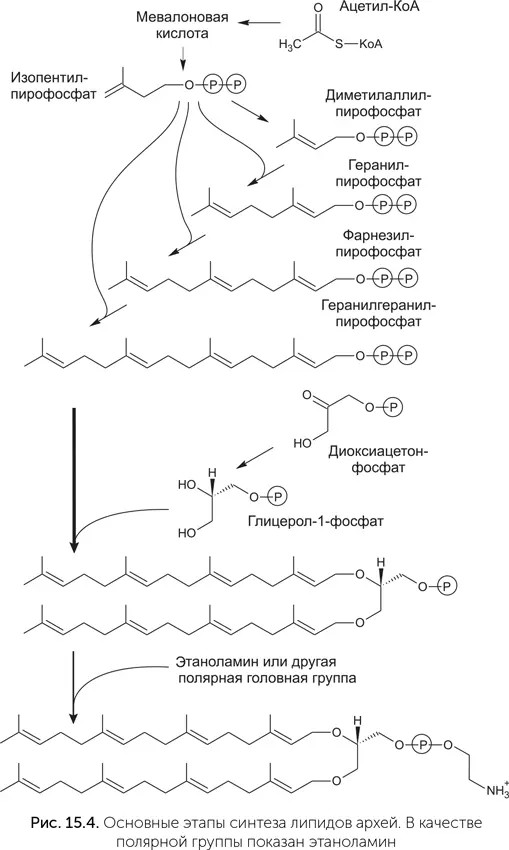
Глицерол-фосфат получается из диоксиацетон-фосфата, и затем к нему присоединяются гидрофобные хвосты. Затем к фосфатной группе прикрепляется нуклеотид цитидин-дифосфат, а на последней стадии он заменяется на полярную головную группу – холин, этаноламин, серин или инозитол.
Новая информация о мембранах LUCA появилась в 2012 году, когда несколькими группами ученых был проведен подробный анализ истории генов всех ферментов биосинтеза всех компонентов липидов бактерий, архей и эукариот (Dibrova et al., 2012; Lombard, Lopez-Garcia, Moreira, 2012).
Краткие результаты исследования приведены в таблице 15.1.
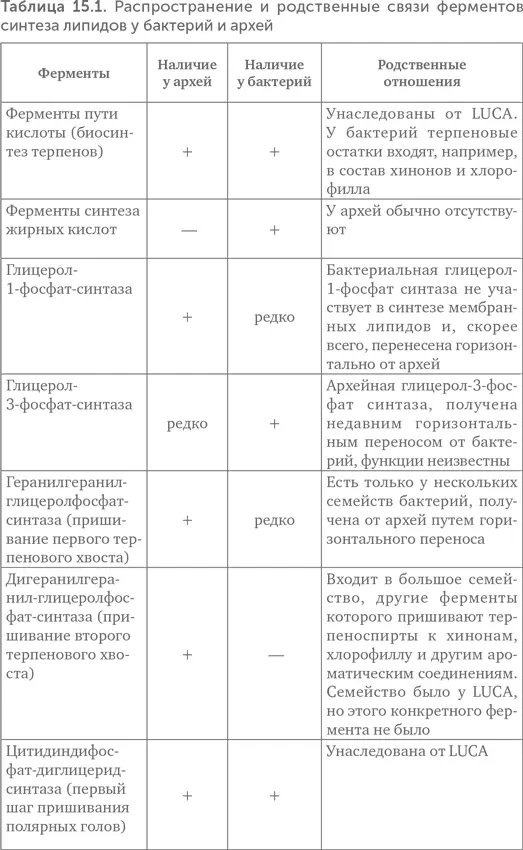
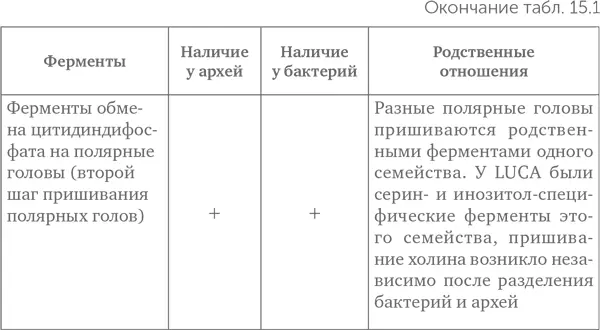
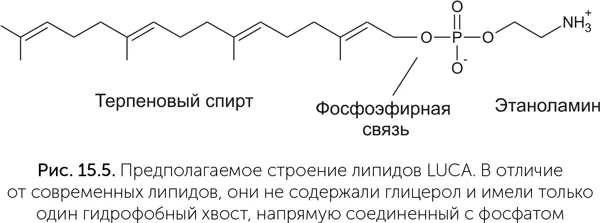
Из таблицы видно, что LUCA мог, во-первых, синтезировать терпеноспирты и, во-вторых, пришивать полярные головы к спиртам. Как синтез глицеролфосфата, так и синтез жирных кислот возникли лишь после разделения линий бактерий и архей. Следовательно, проще всего предположить, что липиды LUCA состояли из одного остатка терпенового спирта, остатка фосфата и полярной группы (серин, этаноламин или инозитол) (рис. 15.5). Подобные липиды были синтезированы искусственно. Образующиеся из них мембраны по сравнению с современными обладают высокой подвижностью, текучестью, хорошо пропускают ионы металлов и малые органические молекулы. Это позволяло древним протоклеткам поглощать готовую органику из внешней среды даже без специальных транспортных белков.
Дополнительным аргументом в пользу большей древности терпенов по сравнению с жирными кислотами в мембранах являются особенности путей синтеза тех и других. Длинные цепи жирных кислот синтезируются одним ферментным комплексом путем последовательного добавления двухуглеродных «кирпичиков» ацетил-КоА по одному. Для получения жирных кислот одинаковой длины (а следовательно, и прочной мембраны) нужны достаточно точные и совершенные ферменты. Терпены же синтезируются из более крупных пятиуглеродных строительных блоков изопентилпирофосфата, которые сначала собираются в десятиуглеродные молекулы геранилпирофосфата, а затем при помощи других ферментов – в более длинные спирты (фарнезол, геранилгераниол, кальдархеол, фитол и др.). Требования к точности и специфичности ферментов при такой крупноблочной сборке гораздо ниже.
Происхождение первых мембранных белков
Не все белки попадают в мембраны при помощи комплекса SRP. Бывают белки, которые синтезируются в одной клетке, а встроиться в мембрану должны в другой, как некоторые бактериальные токсины. Такие белки, чтобы встроиться в мембрану, используют механизм «выворачивания». Для этого белок не должен иметь участков, состоящих только из гидрофобных аминокислот. Неполярные аминокислоты, которые окажутся внутри мембраны, должны чередоваться с полярными примерно 2 аминокислоты через 2. Такой белок может выходить из рибосомы без помощи SRP и достаточно растворим в воде. Чередование «2 через 2» создает альфа-спиральные участки, у которых один бок полярный, а другой неполярный (обсуждалось в главе 13). В водорастворимой форме такой белок сворачивается неполярными участками внутрь, а попадая на мембрану, разворачивается по ней плоско. Затем его альфа-спирали могут опять собраться компактно, но уже гидрофобными участками наружу. Гидрофильные стороны спиралей образуют при этом пору в мембране, через которую могут проходить вода, ионы, полярные молекулы и даже белки и ДНК.
Читать дальшеИнтервал:
Закладка:






