Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Варианты хлорофилльного фотосинтеза
Способностью к фотосинтезу с помощью хлорофиллов или родственных им бактериохлорофиллов обладают пять неродственных групп бактерий: цианобактерии, пурпурные серные бактерии, зеленые серные бактерии (Chlorobi), зеленые несерные бактерии (Chloroflexi) и гелиобактерии (Heliobacteria). Только цианобактерии окисляют воду, выделяют кислород и обладают двумя типами фотосистем в одной клетке. Остальные фотосинтезирующие бактерии не выделяют кислород, более того, их системы фотосинтеза не могут работать в присутствии кислорода, за исключением Chloroflexi.
Серные бактерии, как пурпурные, так и зеленые, для восстановления углекислого газа окисляют сероводород и серу, выделяя сульфаты. Также они могут использовать для фотосинтеза молекулярный водород (выделяя воду) и соли двухвалентного железа (осаждая магнетит, Fe 2O 3). Пурпурные серные бактерии имеют фотосистему второго типа, отдающую электроны на хинон, и фиксируют CO 2в цикле Кальвина. Они особенно многочисленны в сернистых источниках и в озерах, где вода разделена на верхний пресный и нижний соленый слои. В таких озерах отсутствует перемешивание, и нижний соленый слой насыщен сероводородом.
Chloroflexi тоже имеют фотосистему второго типа, подобно пурпурным бактериям, но используют 3-гидроксипропионатный цикл фиксации CO 2(Zarzycky et al., 2009). Их историческое название «зеленые несерные бактерии» неверно, так как они могут использовать для восстановления СО 2сероводород, серу и водород (Tang et al., 2011). Впрочем, самый изученный вид этой группы, Chloroflexus auranticus, в природных условиях обычно не фиксирует СО 2, а потребляет органические кислоты, выделяемые соседями по сообществу, и получает за счет света только АТФ.
Зеленые серные бактерии (Chlorobi) имеют фотосистему первого типа, отдающую электроны на ферредоксин, и используют восстановительный цикл Кребса для фиксации СО 2(Buchanan, Arnon, 1990).
Heliobacteria используют фотосистему первого типа с бактериохлорофиллом g в реакционном центре. Они населяют заболоченные почвы и особенно многочисленны на заливных рисовых полях. Гелиобактерии не способны к автотрофному росту (на СО 2в качестве единственного источника углерода) и нуждаются в готовой органике в виде сахаров или органических кислот, хотя в их геноме закодированы все, кроме одного, ферменты восстановительного цикла Кребса (Tang, Blankenship, 2010). Энергию света они используют для получения АТФ и фиксации азота.
Разные группы фотосинтезирующих бактерий отличаются по предпочитаемой интенсивности света. Фотосистема, настроенная на улавливание слабого света, под ярким полуденным солнцем становится опасной для клетки, вызывая фотохимические повреждения белков и мембран. Многие цианобактерии способны управляться с ярким светом и имеют для этого специальные приспособления: протеинкиназы для быстрой регулировки фотосистем (в течение секунд-минут) путем фосфорилирования белков и маленькие светозащитные белки HLIP (high light induced protein), синтез которых запускается ярким светом. Часть пурпурных бактерий тоже имеют подобные приспособления и переносят яркий свет. Другие фотосинтезирующие бактерии предпочитают укрываться от яркого света под слоем воды или под клетками цианобактерий в бактериальном мате. У Chlorobi и Chloroflexi есть приспособления к очень слабому свету: хлоросомы, зерна из очень плотно упакованных молекул хлорофилла. Благодаря им Chlorobi живут, например, в сероводородном слое Черного моря на глубинах до 100 м.
Эволюция хлорофилльного фотосинтеза
Хлорофилльный фотосинтез используют пять групп бактерий, не родственных между собой. Очевидно, что в распространении генов фотосинтеза большую роль сыграл горизонтальный перенос генов. Это подтверждается и данными по вирусам, заражающим цианобактерий: в их геномах часто встречаются гены компонентов фотосистем. Вирусы могут использовать эти гены для перестройки фотосинтеза зараженной клетки, чтобы быстро получить много энергии для размножения вируса ценой гибели клетки чуть позже. Гены, кодирующие систему фотосинтеза, часто образуют в геноме бактерий компактную группу (супероперон), что должно повышать вероятность переноса всего комплекта.
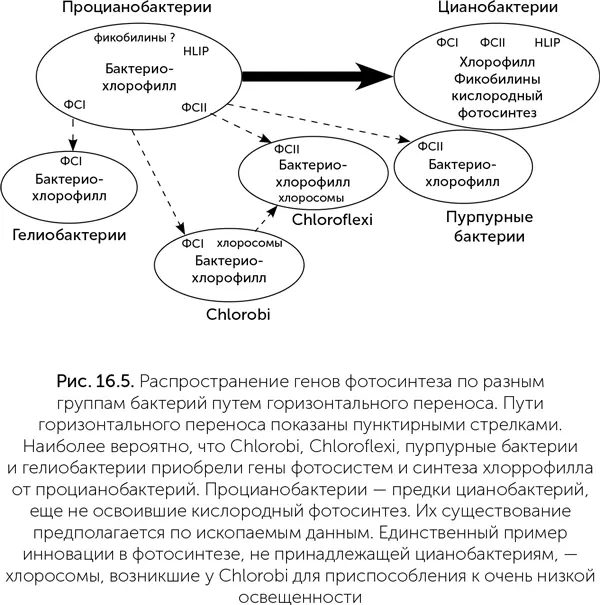
Система фотосинтеза цианобактерий устроена гораздо сложнее, чем у других групп бактерий: только в ней есть два типа фотосистем, дополнительные вспомогательные пигменты (фикобилины) и белки, связывающие их в компактные гранулы (фикобилисомы), а также защитные белки HLIP и протеинкиназы для регуляции фотосистем (рис. 16.5). Если сравнить набор генов, кодирующих детали системы фотосинтеза, у разных групп фотосинтетических бактерий, то получается, что и пурпурные серобактерии, и Chlorobi, и Chloroflexi, и гелиобактерии имеют больше общих генов с цианобактериями, чем друг с другом. Иными словами, горизонтальный перенос генов фотосинтеза происходил в основном либо от цианобактерий, либо к цианобактериям, но не между четырьмя остальными группами (Mulkidjanian et al., 2006). Лишь хлоросомы, служащие для приспособления к очень слабому свету, являются общим признаком Chlorobi и Chloroflexi и, видимо, были перенесены от одной из этих групп бактерий к другой.
Наиболее вероятно, что предки цианобактерий первыми освоили хлорофилльный фотосинтез, и у них же позднее появились две фотосистемы. Другие группы фотосинтезирующих бактерий приобрели способность к фотосинтезу благодаря событиям горизонтального переноса ряда генов от цианобактерий. Скорее всего, эти переносы произошли до того, как цианобактерии освоили кислородный фотосинтез. Фотосистемы Chlorobi и особенно гелиобактерий сохраняют некоторые очень примитивные черты (подробнее об этом – в главе 17).
Геологические следы фотосинтеза и продуктивность древних экосистем
Первые следы фотосинтеза являются одновременно с первыми осадочными породами на Земле, в гренландской формации Исуа возрастом 3,8 млрд лет. Это джеспилиты, или полосатые железные руды (banded iron formations, BIF), а также включения графита со смещенным соотношением изотопов. Полосатые железные руды состоят из тонких (в доли миллиметра) темных слоев магнетита и более светлых слоев кремнезема или карбонатов. Считается, что они отлагались в морях в результате деятельности бактерий, осуществлявших фотосинтез с окислением железа. Магнетит откладывался днем, а карбонатные или кремнеземные слои – ночью. Первые ископаемые, очень похожие на современные нитчатые цианобактерии, появились в осадках Бак Риф (Южная Африка) 3,4 млрд лет назад. Все данные геохимии, однако, свидетельствуют о том, что появление кислорода в атмосфере произошло на 1 млрд лет позже. Изотопное соотношение углерода в осадках Бак Риф указывает на его биологическое происхождение, причем именно через цикл Кальвина. Другие пути фиксации СО 2, такие как ацетил-КоА-путь, более чувствительны к различию в массе изотопов и поэтому сильнее обедняют биологический углерод тяжелым изотопом 13С. Отсутствие серы, сульфатов и магнетита говорит о том, что ни железо, ни сера и сероводород не использовались этими ископаемыми бактериями. Иначе говоря, методом исключения выявлено, что эти древние «процианобактерии» использовали в фотосинтезе водород. Их родство с современными цианобактериями подтверждается и положением в сообществе: они жили в самом верхнем слое бактериального мата на мелководье, а следовательно, были приспособлены к яркому свету.
Читать дальшеИнтервал:
Закладка:






