Михаил Никитин - Происхождение жизни. От туманности до клетки

- Название:Происхождение жизни. От туманности до клетки
- Автор:
- Жанр:
- Издательство:Литагент Альпина
- Год:2016
- Город:Москва
- ISBN:978-5-9614-4350-9
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Михаил Никитин - Происхождение жизни. От туманности до клетки краткое содержание
Происхождение жизни. От туманности до клетки - читать онлайн бесплатно ознакомительный отрывок
Интервал:
Закладка:
Геохимики подсчитали количество водорода, железа и соединений серы, выделяемых из глубин Земли в архейском периоде (3,9–2,5 млрд лет назад). Исходя из этих оценок удалось сравнить продуктивность разных видов бескислородного фотосинтеза. По оценкам Canfield (2006), получается, что в глобальном масштабе вклад железоокисляющего фотосинтеза достигал 90 %. Железоокисляющие фотосинтезирующие бактерии населяли поверхность открытых океанов. Их общая продуктивность могла быть примерно в 10 раз ниже продуктивности современных морских экосистем. Вклад водородного фотосинтеза оценивается примерно в 10 %, а серного – не более 1 %. Однако выход сероводорода из вулканических источников происходит локально, и фотосинтезирующие серобактерии могли образовывать высокопродуктивные «оазисы» в окрестностях вулканов.
Происхождение кислородного фотосинтеза
В течение архейского периода химический состав земной коры и верхней мантии постепенно изменялся: железо погружалось к ядру планеты, вулканическая активность слабела, и продуктивность всех типов фотосинтеза, зависящая от поступления водорода, серы и железа из недр Земли, снижалась. В этих условиях естественный отбор поддержал появление кислородного фотосинтеза, который использует бесконечный источник электронов – воду.
Системы защиты от токсического действия кислорода, необходимые для кислородного фотосинтеза, могли возникнуть даже раньше, чем кислородный фотосинтез. Показано, что на поверхности пирита (FeS 2) при освещении в воде образуется перекись водорода – Н 2О 2. Ионы Fe 3+ – продукт железо-зависимого фотосинтеза – тоже окисляют воду до перекиси водорода при освещении раствора, так что каталаза и другие защитные системы могли возникнуть вскоре после выхода бактерий из обогащенных цинком геотермальных водоемов в моря. Более того, показано, что водоокисляющий комплекс современных цианобактерий может использовать H 2O 2в качестве источника электронов, причем для этого требуется меньше энергии света, чем для окисления воды. Возможно, обезвреживание перекиси водорода было первой функцией примитивного водоокисляющего комплекса. Существуют противоречивые данные о роли H 2O 2в современном кислородном фотосинтезе – в зависимости от изучаемого вида и условий перекись может как подавлять его, так и стимулировать. Некоторым видам цианобактерий для выделения кислорода необходимо, чтобы заметное его количество уже содержалось в среде. Как это объяснить, пока непонятно (Olson, Blankenship, 2004).
Окисление воды – крайне сложная задача. Окислительно-восстановительный потенциал (его сокращенно называют редокс-потенциалом) бактериохлорофилла а (0,55 V) достаточен для окисления водорода, двухвалентного железа, H 2S и серы. Редокс-потенциал хлорофилла а из цианобактерий выше, около 0,82 V. Однако если окислять воду, отбирая электроны по одному, то первая реакция, превращающая воду в гидроксильный радикал и протон, требует огромного редокс-потенциала в 2,75 V, недоступного ни для каких фотосинтетических пигментов. Поэтому необходим промежуточный переносчик электронов, способный окислить сразу две молекулы воды – редокс-потенциал хлорофилла будет достаточен, только если его равномерно делить на четыре электрона.
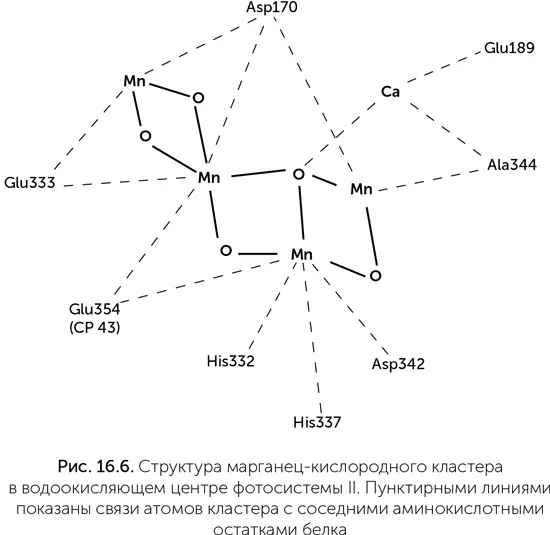
Водоокисляющий комплекс – это связанный с фотосистемой II марганец-кислородный кластер. Он содержит четыре иона марганца, меняющих степень окисления от +3 до +4, кроме того, с ним связано по одному иону кальция и хлора (рис. 16.6). Структура этого кластера напоминает элементарную ячейку таких природных минералов, как рансьеит (CaMn 4O 9× 3H 2O) и других слоистых оксидов марганца, подобно тому как FeS-кластеры окислительно-восстановительных ферментов похожи на ячейку пирита. Существовала гипотеза о происхождении водоокисляющего комплекса от защитного фермента супероксид-дисмутазы, одна из разновидностей которого содержит два иона марганца. Такой примитивный фермент, как предполагалось, мог быть способен к двухэлектронным реакциям – окислению H 2O 2до кислорода. Окисление воды до H 2O 2в такой системе было невозможно из-за недостаточного редокс-потенциала хлорофилла. К сожалению, никакого сходства между последовательностями супероксид-дисмутаз и белков фотосистемы II обнаружить не удалось.
Однако было давно замечено, что выделение кислорода в фотосинтезе сильно стимулируется добавлением бикарбонатов (солей HCO 3-). Добавление изотопно меченого тяжелым кислородом 18O бикарбоната показало, что при освещении в первые секунды выделяется кислород, содержащий только тяжелый изотоп 18О. Это значит, что водоокисляющий центр переключается на окисление бикарбоната, при этом выделяются кислород и углекислый газ. Кроме того, было обнаружено, что бикарбонат участвует в сборке марганцевого кластера. Водорастворимая форма марганца – ионы Mn 2+. В физиологических условиях они находятся в клетке в основном в виде марганец-бикарбонатных комплексов, таких как Mn 2(HCO 3) 2 2+. В процессе сборки марганцевого кластера эти комплексы присоединяются к фотосистеме II. А затем под действием света Mn 2+в составе комплексов окисляются ею до Mn 3+, а бикарбонат – до кислорода и CO 2.
Марганец-бикарбонатные кластеры по своему редокс-потенциалу могут служить донорами электронов для бактериальных фотосистем, при этом происходит окисление Mn 2+до Mn 3+. Это было обнаружено в экспериментах с фотосистемой II пурпурных бактерий, в норме окисляющих железо (Khorobrykh et al, 2007). Из этого можно построить следующую схему происхождения кислородного фотосинтеза (рис. 16.7): сначала, по мере исчерпания железа в океане, предки цианобактерий стали переходить на марганец-бикарбонатные комплексы в качестве источника электронов для фотосинтеза. Затем мутации ФСII привели к тому, что ионы Mn 3+(продукт марганец-окисляющего фотосинтеза) стали задерживаться на белке и принимать участие в его работе. Из них собрался примитивный марганец-кислородный кластер, способный проводить фотоокисление HCO 3–до кислорода и CO 2. Эта реакция требует в полтора раза меньше энергии, чем окисление воды, и возможностей бактериальной ФСII для этого практически достаточно. Так у фотосинтезирующих бактерий появился доступ к новому источнику электронов – бикарбонату. По мере того как запасы марганца в океане были израсходованы и осаждены в виде MnO 2, процианобактерии стали переходить на окисление того, что осталось, т. е. бикарбоната. Марганец, который до того был расходным материалом для фотосинтеза, стал катализатором для использования бикарбонатов, и потребность в нем уменьшилась в тысячи раз.
Читать дальшеИнтервал:
Закладка:






