Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Выявленный у этих гибридов тип летальности указывает, по мнению Стертеванта, на то, что для выживания гибридного потомства необходимо наличие Х-хромосомы D. simulans. Однако даже присутствия этой Х-хромосомы недостаточно для восстановления жизнеспособности, если в цитоплазме D. simulans присутствует также Х-хромосома D. melanogaster. Ключ к непосредственной причине этой летальности, а также подтверждение правильности гипотезы Стертеванта были получены в более поздних исследованиях этого явления. Дьюрика и Кридер (D. Durica, H. Krider) сумели показать, что в гибридных генотипах один из организаторов ядрышка (место синтеза рибосомной РНК) подавлен, что, очевидно, приводит к ослаблению синтеза белка. Большой интерес представляет доминантный аллель, недавно обнаруженный Такамурой и Ватанабе (Т. Takamura, Т. Watanabe) в природе у D. simulans; его присутствие спасает оба летальных класса при реципрокных межвидовых скрещиваниях. Мы вправе допустить, что этот аутосомный локус 2-й хромосомы ответствен за активацию или инактивацию рибосомных генов у межвидовых гибридов. Далее генетические исследования, проведенные несколько лет назад Мёллером и Понтекорво (H.J. Muller, G. Pontecorvo), позволяют считать, что число генов, контролирующих стерильность гибридов, невелико, но оно больше единицы. Хотя летальные взаимодействия, очевидно, детерминируются одним геном, находящимся во 2-й хромосоме, точное число, по-видимому, разбросанных по всему геному локусов, которые обусловливают стерильность жизнеспособного гибридного потомства, еще предстоит выяснить. Более четкая оценка генетической дифференцировки между двумя видами-двойниками дана в конце настоящей главы.
Последний тип постзиготической изоляции называют разрушением гибридов, или гибридным дисгенезом. При такой изоляции гибридные индивидуумы образуются, и они плодовиты; однако у потомков этих гибридных индивидуумов наблюдаются разнообразные нарушения развития: от летальности до пониженной жизнеспособности и стерильности. Примером этого явления служит скрещивание между Zea mays и Ζ. mexicana. Мангельсдорф (Mangelsdorf) получил от этого скрещивания гибридные растения, а затем скрещивал их с растениями Z. mays родительской линии. Потомки от этого скрещивания оказались высоко «мутабильными» и дали в свою очередь потомство с дефектами, затрагивавшими эндосперм, высоту стебля и другие признаки, от которых зависит мощность растений. Подобного рода эффект наблюдается не только у растений; недавно он был обнаружен и проанализирован у Drosophila melanogaster. В этом случае проявления дисгенеза можно видеть у гибридов от скрещиваний между мухами диких линий, выловленными в природе, и лабораторными линиями, которые были изолированы от диких линий в течение всего нескольких лет. Результаты дисгенеза проявляются в виде разрывов хромосом и их элиминации в процессе митотических клеточных делений у развивающихся личинок и в высокой частоте мутаций. Как показал генетический анализ этого процесса, он контролируется одним или несколькими генами и, возможно, связан с перемещением (транспозицией) небольших участков «кочующей» (мобильной) ДНК. Поэтому создается впечатление, что «разрушение» гибридов вызывается генами, влияющими на сохранение генома. В самом деле, возможно, что именно подобного рода механизм вызывает летальность гибридов, наблюдаемую в приведенном выше примере с иглокожими. Наконец, необходимо подчеркнуть, что репродуктивная изоляция редко бывает вызвана одной-единственной причиной; как в примере с гибридами между D. melanogaster и D. simulans, она может быть обусловлена несколькими факторами, блокирующими обмен генами. Некоторые из них могут быть пре-, а другие постзиготическими по своему действию. Конкретная комбинация факторов и природа механизма зависят от участвующих в гибридизации видов, от особенностей их поведения и физиологии размножения.
Механизмы изоляции генофондов сами по себе ничего не говорят о том, как первоначально возникла изоляция. Буш (Buch) обрисовал четыре возможных типа видообразования, которые если и имели место, то привели бы к кладогенезу. Эти четыре типа схематически представлены на рис. 3-6.
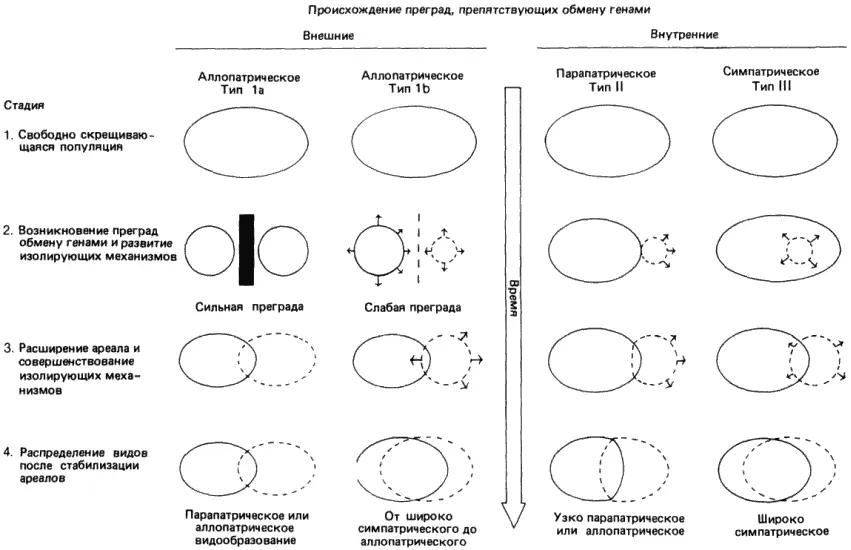
Рис. 3-6.Типы видообразования у животных (Bush, 1975).
Первый из них (тип 1а) обычно называют классическим, или аллопатрическим видообразованием. В исходном состоянии популяция представляет собой обширную панмиктическую (свободно скрещивающуюся) популяцию. Какое-либо изменение географии или климата разбивает эту большую популяцию на две отдельные части и при помощи той или иной физической преграды исключает возможность обмена генами между ними. Под действием отбора или просто в результате случайного дрейфа генетических элементов эти две некогда идентичные популяции превращаются в две генетически обособленные группы. Это изменение происходит не в результате одновременного изменения всех членов данной субпопуляции, а путем медленного замещения первоначальной генотипической конституции исходной популяции, происходящего за счет перераспределения генетического материала. Если физическая изоляция сохраняется на протяжении достаточно длительного времени, то возникает некоторый уровень генетической дивергенции, так что две новые популяции оказываются изолированными при помощи одного или нескольких внутренних изолирующих механизмов, которые мы уже обсуждали. Если на этой стадии устранить внешнюю преграду, то между такими прежде идентичными популяциями обмен генами происходить не будет. Таким образом, согласно определению, можно считать, что возникли два отдельных вида. Если (одновременно с репродуктивной изоляцией) у этих двух популяций появятся и другие признаки, в силу которых они станут занимать разные ниши, то они могли бы совместно обитать в одной и той же географической области (стать симпатрическими в результате миграции). В противном случае между ними может начаться конкуренция, которая приведет к вымиранию одного из двух видов.
Рис. 3-6 содержит еще одно уточнение, касающееся этого способа. Если репродуктивная изоляция несовершенна (особенно в тех случаях, когда возникает только постзиготическая изоляция), то возможен отбор, направленный на создание дополнительных преград. Существование одной лишь постзиготической изоляции привело бы к большой потере гамет для обеих популяций в любой точке их перекрывания, а любая особенность поведения, ведущая к презиготической изоляции, устранит эту потерю и будет подхвачена отбором. Следует указать, что видообразование такого типа протекает медленно.
Читать дальшеИнтервал:
Закладка:









