Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Карсон и Клейтон (F. Clayton) в течение нескольких лет изучали политенные и метафазные хромосомы видов Drosophilidae, эндемичных для Гавайских островов. Они обнаружили, что преобладающее большинство изученных до сих пор видов имеют одинаковый основной метафазный кариотип - пять палочковидных хромосом и одну точечную, а диплоидное число хромосом равно у них 12. Результаты изучения политенных хромосом, позволяющих гораздо детальнее рассмотреть морфологию хромосом, не подтверждают этот кажущийся консерватизм, так как выявляют большое число хромосомных инверсий. Интересно, что эти инверсии представляют собой не просто случайные перестройки хромосом, возникшие у разных видов, а образуют определенные группы и подгруппы; т.е. если произвольно принять последовательность хромосомных дисков у какого-либо вида за стандартную, то другие виды можно сравнивать с этим стандартом, выясняя, по скольким инверсиям они различаются. По мере сопоставления все большего числа «нестандартных» видов со стандартом в наблюдаемых различиях начинает выявляться определенная упорядоченность и возникает возможность построить некую филогению, допустив, например, что если вид А отличается от стандарта группой из пяти или шести инверсий, а вид В - этими же пятью-шестью и вдобавок еще тремя, то представляется вероятным, что вид В находится в более отдаленном родстве со стандартом, чем вид А. Именно таким способом Карсон сумел построить филогенетическое древо для более чем 100 видов гавайских Drosophilidae.
Небольшой участок этого основанного на хромосомных различиях древа представлен на рис. 3-7; он соответствует так называемой группе planitibia этих дрозофил. На кладограмме показаны хромосомные взаимосвязи для 8 из 17 описанных видов этой группы. Если сопоставить приведенную кладограмму с распределением этих 8 видов по разным островам, то выявляется интересная закономерность, позволяющая ввести в созданную схему возможную временную компоненту. Drosophila picticornis встречается только на Кауаи - самом древнем из всех островов, и, судя по структуре его хромосом, этот вид связан с D. attigua и D. primaeva более близким родством, чем другие члены группы planitibia. Считается, что эти два вида относятся к числу самых давних обитателей Гавайских островов и наиболее близки к материковому виду, который, согласно предположениям, первым заселил эти острова и положил начало всей линии гавайских представителей Drosophilidae. На острове Оаху обитают два вида - D. substenoptera и D. hemipeza. Первый из них ближе к D. picticornis и, вероятно, произошел от какого-то сходного с D. picticornis предка, который колонизировал остров Оаху вскоре после его образования, что произошло спустя примерно 2 млн. лет после возникновения острова Кауаи. Судя по морфологии его хромосом, вид D. hemipeza, по-видимому, не связан прямым родством ни с D. picticornis, ни с D. substenoptera, а происходит от какого-то предка, который был сходен с D. neopicta и вторично мигрировал с острова Молокаи обратно на Оаху. Эта гипотеза совместима с наличием у D. hemipeza группы инверсий, сходных с инверсиями, имеющимися у D. neopicta, а также двух других инверсий в Х-хромосоме, одна из которых встречается только у D. hemipeza. Прямое выведение D. hemipeza от D. substenoptera нарушило бы принцип экономии мышления, принятый при построении филогенетического древа. Кроме того, этот аспект филогении хромосом подтверждается биохимическими данными, полученными при анализе полиморфизма по ферментам у этих видов, которые мы приведем чуть ниже.
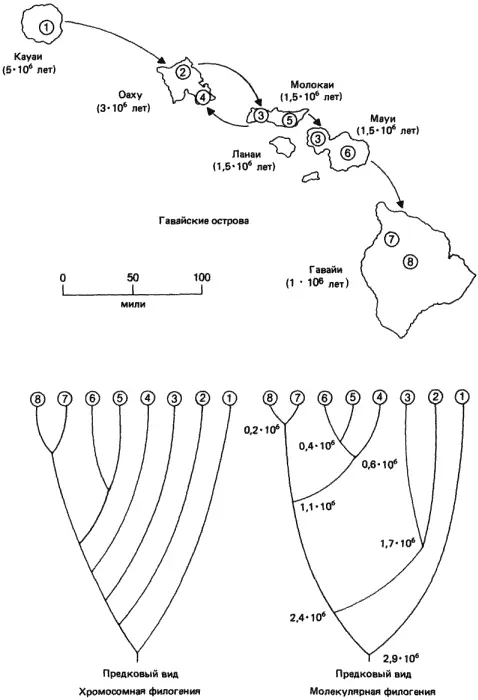
Рис. 3-7.Карта основных островов Гавайского архипелага. Под названием каждого острова в скобках указано время его возникновения (судя по возрасту слагающих его вулканических пород). Стрелками между островами показано направление миграций и случаи основания новых популяций, предположительно имевшие место в процессе эволюции гавайских Drosophilidae. Под картой помещены две дендрограммы, изображающие филогению части группы plantibia. Левая дендрограмма основана на данных об инверсиях, наблюдаемых в политенных хромосомах этих видов. Правая дендрограмма построена по данным вычислений, в основе которых лежит сходство электрофоретической подвижности ферментов, кодируемых 10 разными локусами (сами коэффициенты сходства представлены в табл. 3-2). Значения времени, указанные в каждой точке дивергенции, основаны на определяемой по молекулярным часам скорости замещения аминокислот в исследованных ферментах. Для установления средней скорости этого процесса использовали время возникновения разных островов. Филогении восьми видов, созданные этими двумя независимыми способами, почти идентичны; обе они соответствуют концепции, согласно которой мухи мигрировали с острова на остров, основывая в период заселения островов новые виды. Цифрами в кружках и на дендрограммах обозначены локализация и филогенетические взаимоотношения следующих видов: 1) D. picticornis; 2) D. substenoptera; 3) D. neopicta; 4) D. hemipeza; 5) D. differens; 6) D. planitibia; 7) D. silvestris; 8) D. heteroneura (Johnson et al., 1975).
На острове Молокаи обитает также D. differens, а на соседнем острове Мауи, когда-то соединявшемся с Молокаи, - вид D. planitibia, давший название всей группе. Оба этих вида идентичны по структуре политенных хромосом, и тем не менее они изолированы (как репродуктивно, так и географически). Поскольку D. neopicta обитает на обоих островах, представляется вероятным, что либо D. differens, либо D. planitibia произошли в процессе видообразования от D. neopicta. Какой из этих двух видов, идентичных по своим хромосомам, дал начало другому - менее ясно; нет данных, которые подтверждали бы, что D. differens дал начало D. planitibia, мигрировав с острова Молокаи на Мауи и колонизировав последний, или же что произошло обратное.
Последняя пара изучавшихся видов, D. silvestris и D. heteroneura, встречаются только на самом большом и самом молодом острове Гавайи. Судя по структуре хромосом этих двух видов, которая у них идентична (так же, как у D. differens и D. planitibia), они могли возникнуть в результате колонизации Гавайи каким-то предком, сходным с D. planitibia. Два вида, обитающие на большом острове, существуют почти симпатрически, и тем не менее в природных условиях они редко скрещиваются между собой. В лаборатории, однако, их удается скрестить, получая жизнеспособное и плодовитое потомство. Это, казалось бы, бросает вызов нашему первоначальному определению вида. И все же морфологические различия между этими видами, которые будут описаны в дальнейшем, а также их изоляцию в природе следует рассматривать как указание на то, что их разделение произошло лишь совсем недавно.
Читать дальшеИнтервал:
Закладка:









