Рудольф Рэфф - Эмбрионы, гены и эволюция

- Название:Эмбрионы, гены и эволюция
- Автор:
- Жанр:
- Издательство:Мир
- Год:1986
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Рудольф Рэфф - Эмбрионы, гены и эволюция краткое содержание
В книге американских авторов излагаются факты и идеи о связи генетики, эмбриологии и эволюции. Основное внимание уделено представлению о том, что эволюция идет по преимуществу путем отбора значительных перестроек морфологии, обусловленных мутациями регуляторных генов.
Для специалистов по молекулярной биологии, эмбриологов, генетиков, эволюционистов, для студентов и преподавателей биологических факультетов.
Эмбрионы, гены и эволюция - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Примеры с хрусталиком и поджелудочной железой, а также с хвостовыми мышцами оболочника негативны в том смысле, что диссоциация достигается только в сочетании с утратой некой структуры. Что же касается проблемы приобретения новых структур или функций в процессе эволюции, то здесь было бы более желательным иметь возможность располагать такими примерами диссоциации, в которых цитодифференцировка сочетается с преобразованием структуры. Такие примеры известны, причем, вероятно, наилучший из них описан в работе Сакакура (Sakakura) и его сотрудников. Эти авторы выращивали в культуре эпителий млечной железы 14-дневного мышиного зародыша, комбинируя его с мезенхимой из зачатков млечной или слюнной железы. Затем такие рекомбинированные зачатки выращивали в теле самок сингенных мышей. Как показывают результаты, представленные на рис. 5-5, при объединении эпителия зачатка млечной железы с мезенхимой зачатка той же железы наблюдался морфогенез, типичный для млечной железы; если же такой эпителий объединяли с мезенхимой зачатка слюнной железы, то морфогенез шел по пути, характерному для слюнной железы. Однако, несмотря на свою морфологию слюнной железы, эти последние рекомбинанты в биохимическом отношении вели себя как ткань млечной железы. Так, если самка-хозяин находилась в периоде лактации, то у рекомбинантов наблюдалась пролиферация альвеол с расширенными просветами, синтезировались большие количества типичного белка молока, α-лактальбумина, и альвеолы наполнялись молоком. Морфогенез эпителия млечной железы направлялся индукционными сигналами, исходящими от мезенхимы слюнной железы, но его цитодифференцировка протекала автономно и была, возможно, уже детерминирована отдельным актом индукции, который осуществлялся еще до извлечения зачатка млечной железы из зародыша.
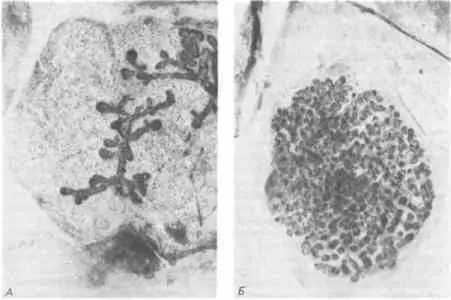
Рис. 5-5.Морфогенез, определяемый индукционными влияниями мезенхимы, при совместном выращивании эпителия и мезенхимы. А. Типичное для млечной железы моноподиальное строение, возникающее при совместном выращивании эпителия и мезенхимы из зачатков млечной железы. Б. Характерное для слюнной железы ветвистое строение, возникающее при совместном выращивании эпителия из зачатка млечной железы и мезенхимы из зачатка слюнной железы; несмотря на морфологию, характерную для слюнной железы, цитодифференцировка эпителия протекает по типу млечной железы (Sakakura, Nishizuka, Dawe, 1976).
О зубах курицы и перьях ящерицы
Если эволюционные изменения в самом деле возникли в результате диссоциации индукционных событий, то можно ожидать, что остатки прежних взаимосвязей должны сохраняться, что регуляторные гены, не выявляющиеся таким образом, как прежде, все еще присутствуют. Подобное ожидание вполне обоснованно, как показывает рис. 5-6, на котором О. Мангольд (Mangold) схематически изобразил некоторые из основных индукционных событий в голове и туловище зародышей амфибий. В этой схеме подчеркивается общее заключение, уже выведенное из работ Джекобсона, о сложности индукционных взаимодействий, которые, как правило, принадлежат к типу переплетающихся или каскадных взаимодействий. Применительно к эволюции это означает, что индукционные взаимодействия устойчивы к существенным модификациям и что в случае возникновения достаточно крупных изменений они обычно не выходят за рамки характерных для данной группы. Коренные изменения, с которыми сопряжено возникновение некоторых новых групп, таких как членистоногие (см. гл. 4), иногда имеют место, но это случается редко. Другое следствие интегрированности морфогенетических путей, о котором часто упоминают, состоит в том, что эволюционные изменения, относящиеся к поздним стадиям развития, воспринимаются легче, чем ранние изменения, просто потому, что поздние изменения требуют меньше соответствующих изменений в затрагиваемых ими каскадных процессах. Вряд ли, однако, можно считать, что изменения в онтогенезе возникают исключительно таким образом; если бы это был единственный способ, то что-то очень близкое к геккелевской рекапитуляции действительно играло бы универсальную роль. На самом же деле изменения возникают на всех стадиях развития, так что хотя картины взаимодействий, наблюдаемые у амфибий, и можно считать своего рода архетипом для развития позвоночных, среди последних нельзя найти двух видов, развитие которых протекало бы совершенно одинаково, пусть даже на самых ранних стадиях. Раннее сходство и все возрастающее в дальнейшем обособление зародышей родственных организмов, о которых говорится в законах Бэра, не следует понимать чересчур буквально. Развивающиеся млекопитающие ни на какой стадии развития не бывают идентичны зародышам рыбы или ящерицы: по своим геномам и характеру развития они очень далеко ушли от своих предков. Но тем не менее некое исконное сходство между ними сохранилось. А произошло следующее: множественные индукционные взаимодействия, описанные на примерах индукции хрусталика, носа и уха, создали достаточно высокий уровень морфогенетического гомеостаза, так что изменения могут восприниматься даже на ранних стадиях общего процесса развития, не нарушая его интегрированности.
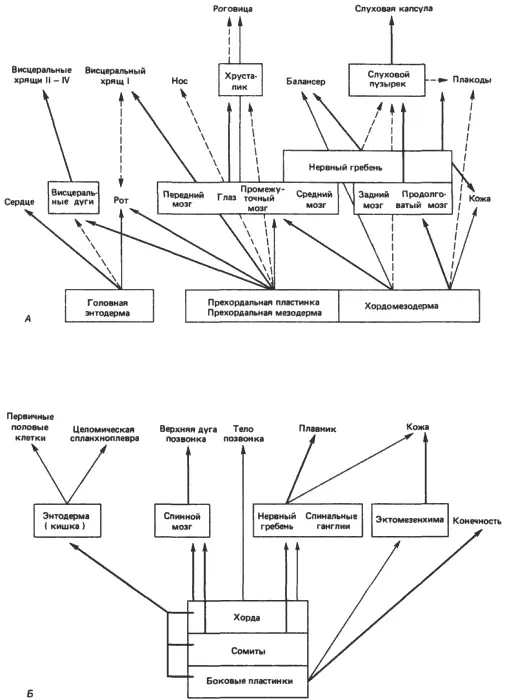
Рис. 5-6.Индукционные взаимодействия и каскадные процессы в развитии амфибий. А. Индукционные процессы в развитии головы. Б. Индукционные процессы в развитии туловища. Толщина стрелок отражает относительную интенсивность индукции (Mangold, 1961).
Существование общего консервативного плана развития позволяет считать, что эмбриональные структуры могут сохраняться даже после того, как они утратили свою прежнюю основную функцию, потому что они все еще служат связующими звеньями в каскаде морфогенетических взаимодействий, присущих прежней функции. Таким же образом могут сохраняться и регуляторные системы. Доказательством сохранения прежних регуляторных генов могло бы служить воскрешение отвергнутого в прошлом пути развития при нарушении существующего типа развития в результате мутации или экспериментального воздействия. Известны примеры того и другого. Восстановление старых типов развития в результате мутаций подробно рассматривается в гл. 8 и 9. Экспериментальный анализ трех примеров эволюционных изменений позволил обнаружить модификации в системах взаимодействия между тканями, при которых на прежние механизмы регуляции налагались бы новые, но без утраты генетической информации, имевшейся у предков. Два из этих примеров относятся к модификациям, имевшим место в эволюции птиц; третий касается изменений в покровных тканях рептилий, птиц и млекопитающих.
Читать дальшеИнтервал:
Закладка:









