Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
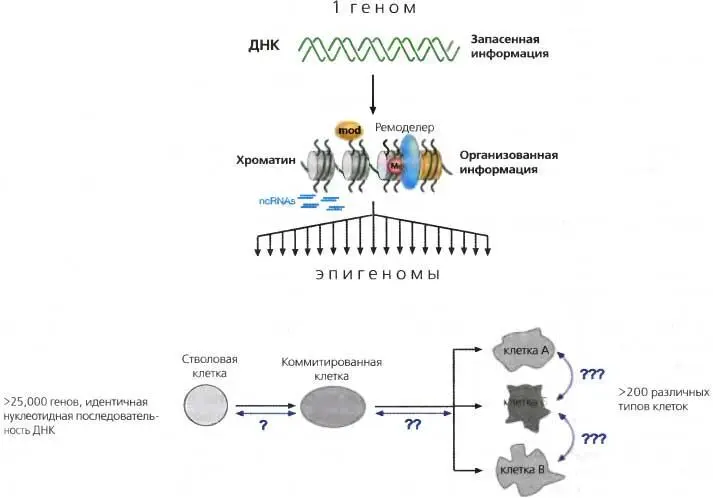
Рис. 3.4.ДНК vs. хроматин
Геном: инвариантная нуклеотидная последовательность ДНК ( зеленая двойная спираль ) особи. Эпигеном: общий состав хроматина, индексирующий весь геном в любой данной клетке. Он варьирует в зависимости от типа клетки и реакции на внутренние и внешние сигналы, которые он получает. ( Нижняя часть рисунка ) эпигеномная диверсификация у многоклеточных организмов происходит в ходе развития по мере того, как дифференцировка прогрессирует от единичной стволовой клетки (оплодотворенный эмбрион) к более коммитированным клеткам. Реверсия дифференцировки или трансдифференцировка ( голубые линии ) требует репрограммирования эпигенома клетки
Фенотипические изменения, происходящие в ряду клеточных поколений в ходе развития многоклеточного организма, были описаны Уодцингтоном как «эпигенетический ландшафт» (Waddington, 1957). Тем не менее, весь спектр клеток, от стволовых до полностью дифференцированных, обладает идентичными нуклеотидными последовательностями ДНК, но заметно различается по профилю генов, которые реально экспрессируются этими клетками. Исходя из этого, позднее пришли к определению эпигенетики как «ядерной наследственности, которая не основывается на различиях в нуклеотидной последовательности ДНК» (Holliday, 1994).
Со времени открытия двойной спирали ДНК и ранних трактовок эпигенетики наши знания об эпигенетическом контроле и лежащих в его основе механизмах существенно возросли, заставляя некоторых описывать эти знания в таких более «возвышенных» терминах, как «область науки», а не просто «феномены» (см Wolfe and Matzke, 1999; Roloff and Nuber, 2005; глава 1). За последнее десятилетие значительный прогресс был достигнут в отношении многих семейств энзимов, активно модифицирующих хроматин (см. ниже). Таким образом, используя современную терминологию, эпигенетику можно в молекулярном (механистическом) плане определить как «сумму изменений в хроматиновой матрице, которые в совокупности устанавливают и воспроизводят различные паттерны экспрессии генов (транскрипции) и сайленсинга на основе одного и того же генома».
4. Хроматиновая матрица
Нуклеосома является фундаментальной повторяющейся единицей хроматина (Komberg, 1974). С одной стороны, эта базовая единица хроматина состоит из белкового октамера, содержащего по две молекулы каждого канонического (или корового) гистона (Н2А, Н2В, H3 и Н4), вокруг которого накручены 147 п.н. ДНК. Детальные межмолекулярные взаимодействия между коровыми гистонами и ДНК были определены в выдающихся исследованиях, приведших к получению рентгеновской картины (с атомным, 2.8 Е, разрешением) нуклеосомы, собранной из рекомбинантных частей (рис. 3.5) (Lugeret al., 1997). Картины мононуклеосом, а также возникающих структур более высокого порядка (тетрануклеосом), имеющие более высокое разрешение (Schalch et al., 2005), продолжают привлекать наше внимание, обещая помочь в объяснении физиологически важного субстрата, на котором развертывается действие если не всего, то большей части ремоделинга хроматина и механизма транскрипции
Коровые гистоновые белки, составляющие нуклеосому, являются очень небольшими и сильно основными. Они состоят из глобулярного домена и гибких (относительно неструктурированных) «гистоновых хвостов», торчащих с поверхности нуклеосомы (рис. 3.5). Судя по аминокислотной последовательности, гистоновые белки крайне консервативны в диапазоне от дрожжей до человека. Такая высокая степень консервативности подкрепляет общий взгляд, согласно которому эти белки, даже неструктурированные хвостовые домены, вероятно выполняют критичные функции. Хвосты, особенно хвосты гистонов H3 и Н4, в действительности содержат важный ключ к изменчивости нуклеосом (и, отсюда, хроматина), поскольку многие из остатков являются объектами экстенсивных посттрансляционных модификаций (см. стандартную номенклатуру, используемую в этом руководстве, на обратной стороне задней части переплета и список известных модификаций гистонов в Приложении 2).
Ацетилирование и метилирование коровых гистонов, особенно H3 и Н4, были одними из первых описанных ковалентных модификаций, и долгое время предполагалось, что они коррелируют с положительными и отрицательными изменениями в транскрипционной активности. Со времени пионерских работ Олфри и сотрудников (Allfrey et al., 1964) были идентифицированы и охарактеризованы многие типы ковалентных модификаций гистонов; в их числе — фосфорилирование, убиквитинирование, сумоилирование, АДФ-рибозилирование, биотинилирование гистонов, изомеризация пролина и другие вероятные типы, ожидающие своего описания (Vaquero et al., 2003). Эти модификации происходят в специфических сайтах и остатках, некоторые из них изображены на рис. 3.6 и перечислены в Приложении 2. Эту ковалентную маркировку катализируют специфические ферменты и ферментные комплексы, часть которых описывается далее в этом обзоре и в отдельных главах. Поскольку в ближайшие годы эти перечни будут продолжать расти, нашим намерением было упомянуть лишь отдельные метки и энзимы, которые могли бы проиллюстрировать то, что, как нам кажется, является общими понятиями и принципами.
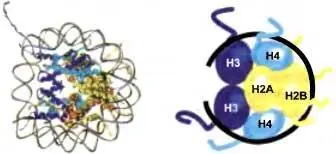
Рис. 3.5.Структура нуклеосомы
(Слева) Модель нуклеосомы с разрешением 2.8 Е. ( Справа ) Схематическое представление организации гистонов внутри октамерного кора, вокруг которого обернута ДНК ( черная линия ). Образование нуклеосомы происходит сперва через откладку на ДНК тетрамера H3/Н4. а затем двух наборов димеров Н2А/Н2В. Неструктурированные аминотерминальные гистоновые хвосты выпячиваются из нуклеосомного кора, состоящего из структурированных глобулярных доменов восьми гистоновых белков
В определенных районах хроматина нуклеосомы могут содержать вариантные гистоновые белки вместо какого-нибудь корового (канонического) гистона. Текущие исследования показывают, что это различие в составе способствует выполнению специализированных функций этими маркированными районами хромосом. В настоящее время известны вариантные белки для коровых гистонов Н2А и H3, но не известно ни одного для гистонов Н2В и Н4. Мы подозреваем, что варианты гистонов, хотя они нередко являются минорными в смысле их количества и потому более трудными для исследования, весьма богаты в отношении содержащейся в них информации и играют весьма существенную роль в отношении их вклада в эпигенетическое регулирование (детали см в разделе 8 и главе 13).
Читать дальшеИнтервал:
Закладка:



