Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
5. Более высокие уровни организации хроматина
Хроматин, этот состоящий из ДНК и нуклеосом полимер, является динамической молекулой, существующей во многих различных конфигурациях. Ранее в течение длительного времени хроматин разделяли на эухроматин и гетерохроматин, исходя из картины окрашивания ядра красителями, которые цитологи использовали для визуализации ДНК. Эухроматин представляет собой деконденсированный хроматин, хотя он может быть активным или неактивным в отношении транскрипции. Гетерохроматин можно определить в широком смысле как высококомпактизированный и «молчащий» хроматин. Он может существовать как постоянно «молчащий» хроматин (конститутивный гетерохроматин), где гены организма лишь изредка экспрессируются в клетках любого типа, или как хроматин, репрессированный в некоторых клетках в ходе специфического клеточного цикла или на специфической стадии развития (факультативный гетерохроматин) Таким образом, имеется спектр состояний хроматина, и накопленная за многие годы литература позволяет предполагать, что хроматин является высоко динамичной структурой, склонной к ремоделингу и реструктурированию по мере получения физиологически релевантных сигналов, поступающих по «идущим вверх» (upstream) сигнальным путям. Однако лишь недавно достигнут значительный прогресс в раскрытии молекулярных механизмов, управляющих этими процессами ремоделинга.
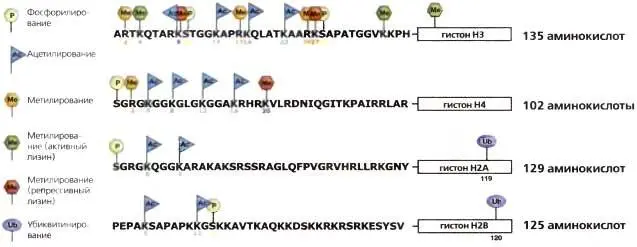
Рис. 3.6.Сайты, по которым происходят модификации гистоновых «хвостов»
Аминотерминальные «хвосты» гистонов составляют четверть массы нуклеосомы. На них находится огромное большинство известных сайтов, по которым происходит ковалентная модификация и которые изображены на рисунке. Модификации происходят также и в глобулярном домене ( заключено в прямоугольник ) некоторые из них указаны на рисунке. В целом в число активных меток входят ацетилирование ( бирюзовый флажок Ас ), метилирование аргинина (желтый шестиугольник Ме) и метилирование некоторых лизинов, таких как H3К4 и H3K36 ( зеленый шестиугольник Ме ). Функция H3К79 в глобулярном домене — антисайленсинг. В число репрессивных меток входят H3К9, H3К27 и H4K20 ( красный шестиугольник Ме ). Зеленый цвет — активная метка, красный — репрессивная метка
Фигурирующая в учебниках 11-нанометровая матрица типа «бусинок на нити» представляет собой активную и в основном «развернутую» интерфазную конфигурацию, в которой ДНК периодически оборачивается вокруг повторяющихся единиц — нуклеосом (рис. 3.7). Однако эта хроматиновая фибрилла не всегда составлена из нуклеосом, расположенных на регулярном расстоянии друг от друга. Нуклеосомы могут быть упакованы нерегулярно и могут сворачиваться в структуры более высокого порядка, анализ которых при атомном разрешении еще только начинается (Khorasanizadeh, 2004). Характерные конформации хроматина более высокого порядка возникают в разных районах генома во время спецификации судьбы клеток или на разных стадиях клеточного цикла (интерфазный vs. митотический хроматин).
Расположение нуклеосом на 11-нанометровой матрице может быть изменено в результате cis -эффектов и trans -эффектов ковалентно модифицированных «хвостов» гистонов (рис. 3.8). Cis -эффекты возникают в результате изменений в физических свойствах модифицированных хвостов гистонов, таких как модуляции в электростатическом заряде или в структуре «хвостов», которые, в свою очередь, изменяют межнуклеосомные контакты. Хорошо известным примером может служить ацетилирование гистонов, которое, как давно подозревали, нейтрализует положительные заряды сильно основных гистоновых «хвостов», порождая в результате локальное растяжение хроматиновой фибриллы и тем самым обеспечивая лучший доступ транскрипционной машины к двойной спирали ДНК. Фосфорилирование, благодаря добавлению суммарного отрицательного заряда, может создавать «очажки зарядов» [charge patches] (Dou and Gorovsky, 2000), которые, как полагают, изменяют упаковку нуклеосом или экспонируют аминоконцы гистонов, изменяя у хроматинового полимера состояние сворачивания более высокого порядка (Wei et al., 1999; Nowak and Corces, 2004). Считается, что во многом таким же образом линкерные гистоны (Н1) стимулируют упаковку фибрилл более высокого порядка, экранируя отрицательный заряд линкерной ДНК между соседними нуклеосомами (Thomas, 1999; Khochbin, 2001; Harvey and Downs, 2004; Kimmins and Sassone-Corsi, 2005). Добавление таких объемистых аддуктов, как убиквитин и АДФ-рибоза, также может индуцировать разное расположение гистоновых «хвостов» и открывать группы нуклеосом. В какой мере гистоновые «хвосты» могут индуцировать компактизацию хроматина посредством механизмов, зависящих от модификаций и не зависящих от них, — еще не ясно.
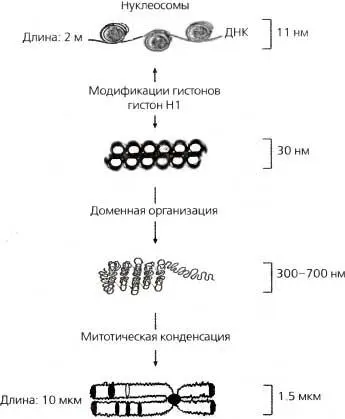
Рис. 3.7.Структурирование хроматина более высоких порядков
11-нанометровая фибрилла представляет собой ДНК, накрученную вокруг нуклеосом 30-нанометровая фибрилла компактизирован а далее во все еще неподтвержденную структуру ( изображена здесь как соленоидная конформация ), включающую линкерный гистон Н1. 300—700-нанометровое волокно представляет собой динамическое «выпетливание» более высокого порядка, которое обнаруживается как в интерфазном, так и в метафазном хроматине. 1,5-микрометровая конденсированная хромосома представляет собой наиболее компакгизированную форму хроматина, которая обнаруживается только во время ядерного деления (митоза или мейоза). Еще не ясно, каким образом картины бэндинга (т. е. G- или R-бэндинга) митотической хромосомы коррелируют с конкретными структурами хроматина

Рис. 3.8.Переходы в хроматиновой матрице (cis/trans)
cis-эффекты : ковалентная модификация остатка гистонового «хвоста» приводит к изменению структуры или заряда, проявляющемуся как изменение в организации хроматина, trans-эффекты : ферментативная модификация остатка гистонового «хвоста» (напр., метилирование H3К9) приводит к возникновению сродства к ассоциированному с хроматином белку (modbinder, напр., НР1). Ассоциация mod binder (или комплексов ассоциированных белков) вызывает изменения в структуре хроматина «вниз по течению». Замещение гистонов: ковалентная модификация гистонов (или другой стимул) может послужить сигналом для замещения какого-то корового гистона вариантным гистоном с помощью комплекса-обменника, осуществляющего ремоделинг нуклеосом
Читать дальшеИнтервал:
Закладка:



