Владимир Цыркунов - Клещевые инфекции в Беларуси

- Название:Клещевые инфекции в Беларуси
- Автор:
- Жанр:
- Издательство:ГрГМУ
- Год:2009
- Город:Гродно
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Владимир Цыркунов - Клещевые инфекции в Беларуси краткое содержание
Клещевые инфекции в Беларуси - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
В репликативном цикле геномная РНК является инициатором инфекционного процесса и служит матрицей для создания дочерних копий.
Нуклеотидная последовательность всего генома ВКЭ и полная аминокислотная последовательность полипротеина вируса расшифрованы Плетневым с соавторами. На рис. 1 представлена линейная генетическая карта вируса с указанием кодируемых генов всех структурных и неструктурных белков и концевых нетранслируемых областей генома ВКЭ штамма Софьин.
Немного позже были расшифрованы полная нуклеотидная последовательность генома ВКЭ западного штамма Neudoerfi (Найдорф), дальневосточного штамма 205 и другого вируса из антигенного комплекса ВКЭ – вируса Повассан.
Первым с N–концевой части полипротеина ВКЭ находится нуклеокапсидный Core–белок С. Молекулярная масса – 12108 Da. При сборке вируса он вместе с геномной РНК образуют центральную структурную компоненту нуклеокапсида.
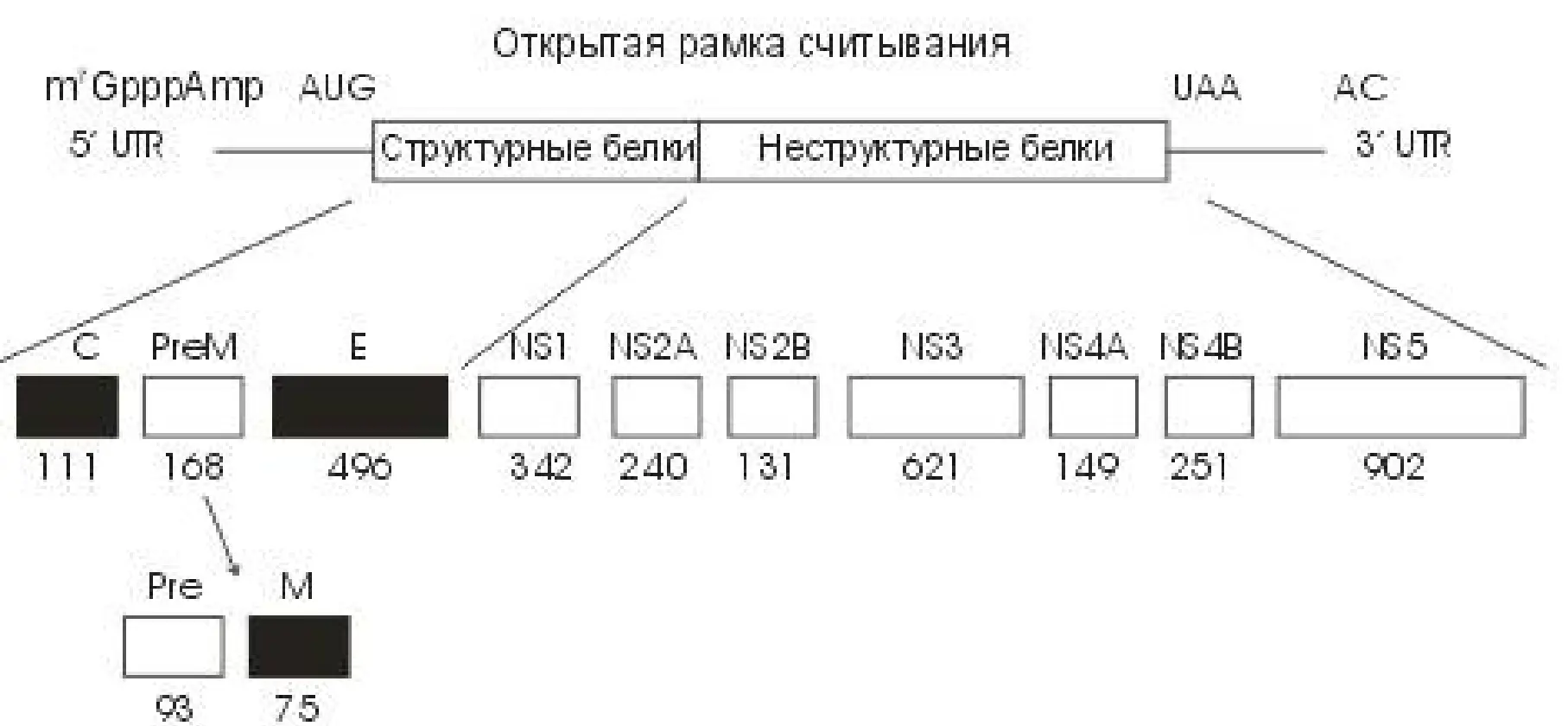
Рис. 1. Структура генома ВКЭ. Расположение концевых нетранслируемых областей, С, PreM (Pre+M), E и NS1–NS5 генов соответствующих структурных и неструктурных белков вируса.
Следующим от N–конца фрагментом полипротеина, отщепляемым протеазами, является гликопротеин ргеМ–полипептид. PreM является только предшественником в синтезе вирионного белка М, его нет в составе зрелого вириона в крови, не удается обнаружить его и в зараженных вирусом клетках. Мембранный белок М, содержащийся в составе оболочки зрелого вируса, представлен 75 остатками аминокислот, имеет молекулярную массу 8209 Da.
Следующим, отщепляемым от предшественника полипептидом является наиболее важный поверхностный вирионный белок Е – главный, биологически наиболее значимый компонент внешней поверхности оболочки вириона. Он играет ключевую роль в процессах сборки вирусной частицы, в связывании вируса с клеточной поверхностью, последующем слиянии вирусной и клеточной мембран, развитии гуморального и клеточного иммунитета. Полипептидная цепь белка Е с молекулярной массой 53680 Da, заканчивается двумя гидрофобными участками, вероятно, служащими трансмембранным «якорем» белка. Считается, что процессинг полипротеина между белком Е и неструктурным белком NS1 осуществляет связанная с мембранами аппарата Гольджи протеиназа.
Современное исследование штаммов вируса КЭ (В.И.Злобин, О. З. Горин) показало, что на территории России обнаружено, пять антигенных вариантов (подтипов) вируса КЭ, имеющих распространение на определенных территориях:
■ Дальневосточный подтип вируса КЭ – Дальний Восток, Восточная и Западная Сибирь, Прибалтика, Калининградская область, Урал.
■ Западный (центрально – европейский) подтип – Центральная Европа.
■ Вергина – Центральная Европа, Удмуртия, Алтай, Ярославская область, Западная Сибирь.
■ Восточносибирский подтип – Восточная Сибирь.
■ Урало–сибирский подтип – Урал, Восточная и Западная Сибирь, Северо–запад Росси, Центральная Азия.
До последнего времени считалось, что на территории Беларуси циркулируют штаммы КЭ западного генотипа, Однако исследованиями, проведенными совместно с В.И. Злобиным, показано, что в Республике возможна циркуляция других генотипов вируса КЭ. Полученные результаты гибридизации показали, 9 штаммов относятся к генотипу 2 (западному) + штамм 256, один штамм 118–КЭ – к генотипу 1 (дальневосточному) + штамм Софьин.
Во внешней среде в составе мозговой суспензии на физиологическом растворе полностью собранная частица ВКЭ сохраняет жизнеспособность при комнатной температуре до 10 дней.
Вирус устойчив в кислой среде, в частности, в составе желудочного сока; вирус в крови или его суспензия в белковом растворе инактивируются при 56°С в течение 30 мин. Кипячение убивает вирус в течение 2–3 мин. В глубоко замороженном виде или лиофилизированном состоянии в вакууме вирус сохраняет жизнеспособность многие годы. Благодаря бислойной липидной природе оболочки, вирус чувствителен к эфиру, растворам лизола, спиртам.
Эпидемиология
Ареал вируса клещевого энцефалита в значительной степени совпадает с видовыми ареалами главных переносчиков I. persulcatus и I. ricinus. Соответственно, вирус клещевого энцефалита встречается на обширной территории от Атлантического до Тихого океана в различных типах лесных экосистем. Инфекция широко распространена по всей лесной и лесостепной умеренной климатической зоне Евразийского континента. Природные очаги имеются во всех странах западной, центральной, восточной и отчасти северной Европы, в частности, на Британских островах, во Франции, Германии, Швеции, Австрии, Чехии, Словакии, Венгрии, Польше, Беларуси, Российской Федерации, Прибалтийских странах.
ВКЭ переносится клещами вида I. persulcatus и I. ricinus. В некоторых очагах КЭ установлена зараженность вирусом других видов клещей: H. concinna, D. silvarum, D. pictus и др. Считается, что еще до 14 видов клещей являются носителями вируса в природе, но их эпидемическая роль невелика. Основным резервуаром вируса в природе являются его главные переносчики, иксодовые клещи. Врожденная инфицированность клещей поддерживает постоянный уровень вирусофорности этих насекомых. Дополнительным природным резервуаром вируса служат прокормители инфицированных клещей: многочисленные грызуны (полевая мышь, бурундук, заяц и др.), другие дикие и домашние животные, птицы. В природе вирус поддерживается постоянной циркуляцией по замкнутой цепи: клещи – животные–прокормители–клещи.
Инфицирование человека для ВКЭ является биологически тупиковым направлением, поскольку вирус дальше не передается к другому организму и выходит из природной циркуляции.
Для КЭ характерна весенне–летняя сезонность заболеваемости, связанная с сезонной активностью переносчиков. В ареале I. persulcatus подъем заболеваемости приходится на весну и первую половину лета, май–июнь месяцы, когда наиболее высока биологическая активность этого вида клещей. Для клещей вида I. ricinus характерен подъем биологической активности дважды за сезон. Инфицирование человека ВКЭ происходит во время кровососания голодных, зараженных вирусом, клещей. Кровососание самки клеща продолжается много дней, и при полном насыщении она увеличивается в весе 80–120 раз. Кровососание самцов длится обычно несколько часов и иногда остается незамеченным. Передача вируса КЭ может происходить в первые минуты присасывания зараженного клеща к человеку. Удаление клеща при сохранении «цементирующей пробки» из содержащей вирус слюны клеща в коже человека, вероятно, полностью не устраняет передачу инфекции. В одном высоковирусофорном клеще может содержаться преизбыточная доза возбудителя инфекции – до 10 10вирусных частиц, из числа которых попадание в кровь человека одной миллионной доли вирусного пула может быть достаточно для развития инфекции. Численность иксодовых клещей в природных очагах на территории Беларуси составляет 2,5–3,5 особей на 1 флаго/км, а циркуляция вируса КЭ установлена в 93 районах республики (С.Е. Яшкова, Н.С. Себут, 2006). На территории Гродненской области численность клещей в природных очагах КЭ составляет 2,5–3,4 особей на 1 флаго/км (А.Р. Хутко с соавт., 2007), при индивидуальной зараженности (вирусофорности) клещей в естественных (диких) природных очагах от 2 до 15%. Уровень вирусофорности отдельного клеща зависит от его возраста и степени «напитанности». Максимальное содержание вируса устанавливается у клещей предельных физиологических возрастов. Низкий уровень вирусофорности у клеща наблюдается в фазе перехода его к активному состоянию. Доля клещей с высоким содержанием вируса среди напитавшихся особей значительно выше, чем у голодных сородичей. На основании этого феномена предполагается возможность ускоренного размножения вируса во время и после питания клеща. Концентрация вируса в I. persulcatus, напитавшихся на больных мышах, непрерывно возрастает, достигая максимума к 40 дню. За этот период количество вируса возрастает в 1000 раз.
Читать дальшеИнтервал:
Закладка:









