Петр Анохин - Проблемы лечебного голодания. Клинико-экспериментальные исследования

- Название:Проблемы лечебного голодания. Клинико-экспериментальные исследования
- Автор:
- Жанр:
- Издательство:Типография №1 Росглавполиграфкомбината Комитета по печати при Совете Министров РСФСР
- Год:1969
- Город:Москва
- ISBN:нет данных
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Петр Анохин - Проблемы лечебного голодания. Клинико-экспериментальные исследования краткое содержание
Представленные в этом сборнике работы дают подробное описание клиники лечебного голодания, нейрофизиологии голода и насыщения, патофизиологии и биохимии голодания. Авторами описано большое количество клинических примеров излечения больных с помощью лечебного голодания от целого ряда психических и соматических заболеваний, в том числе считающихся «неизлечимыми» и трудно поддающимися лечению.
Сборник будет интересен врачам различных специальностей, психиатрам и психотерапевтам, патофизиологам и специалистам других медицинских и биологических наук, студентам высших и средне-специальных медицинских учебных заведений, искателям здорового образа жизни и всем остальным, кто глубоко интересуется лечебным голоданием, включая тех, кто уже имеет личный опыт этой методики, поскольку представленная информация имеет огромную научную ценность для понимания сути процессов, происходящих в процессе лечебного голодания.
Этот сборник вышел в 1969 году, но, несмотря на это, до сих пор распространен миф, что голодание свыше нескольких дней наносит непоправимый вред организму. Поэтому одна из главных задач этого тома — продемонстрировать физиологичность голодания и безвредность больших сроков при этой методике.
Проблемы лечебного голодания. Клинико-экспериментальные исследования - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
Отметка времени 1 сек.
Приступая к настоящей серии экспериментом, мы исходили из следующего предположения. Состояние голода на основе восходящих активирующих влияний подкорковых образований, создает в центральной нервной системе определенное доминантное состояние, сопровождающееся мобилизацией определенных корковых синаптических образований. В этом состоянии любое возбуждение другой биологической модальности, поступающее к коре мозга, должно было неизбежно вступить в какие-то взаимодействия с наличным пищевым возбуждением и, естественно, претерпеть определенные изменения. Особенно отчетливо эти изменения должны быть выражены у соматосензорного вызванного ответа, поскольку формирующее его возбуждение адресуется к тем же передним отделам -коры мозга, куда поступают и возбуждения пищевого центра.
Проведенные опыты показали следующее. У голодных животных, находящихся под уретановым наркозом, соматосензорные вызванные потенциалы в ответ на одиночное раздражение седалищного нерва регистрировались только в ограниченном участке сензомоторной области коры мозга, т. е. в той области, которая является специфической проекционной зоной соматосензорных возбуждений.
Как правило, в этом пункте регистрировались вызванные ответы только с первичноположительной фазой. В ряде опытов у голодных животных соматосензорные вызванные ответы вообще не регистрировались. Характерно, что в то же время и в том же пункте коры вызванные потенциалы, возникающие у голодных животных при раздражении пищевых центров гипоталамуса, выявлялись в коре мозга совершенно отчетливо.
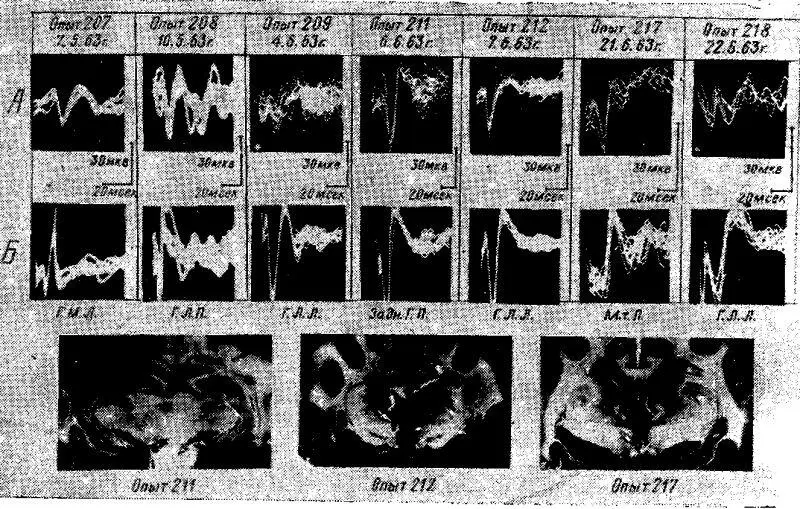
Рис. 12. Вызванные потенциалы в сензомоторной области коры у голодных кошек, находящихся под уретановым наркозом, при раздражении седалищного нерва (А) и различных отделов гипоталамуса 1Б.

Pиc. 13. ЭЭГ (внизу) и карта распространения соматосензорного вызванного потенциала по коре мозга кошки, накормленной перед опытом. Уретановый наркоз.
Обозначения: ЛП — лобная правая, ЛЛ — лобная левая. СМП — сензомоторная правая. СМЛ — сензомоторная левая, ТП — теменная правая, TЛ — теменная левая, ЗП — затылочная правая, 3Л — затылочная левая обл. Отметка времени 1 сек.
Наоборот, у накормленных перед опытом животных соматосензорные вызванные ответы с отчетливо выраженными первичной положительной и отрицательной фазами и нередко с вторичным положительным колебанием регистрировались над обширной областью передних отделов коры мозга.
Аналогичный эффект возрастании амплитуды отрицательной фазы в фокусе максимальной активности и расширения зоны регистрации вызванных ответов наблюдался при введении голодным животным раствора глюкозы или при -коагуляции медиальных отделов гипоталамической области.
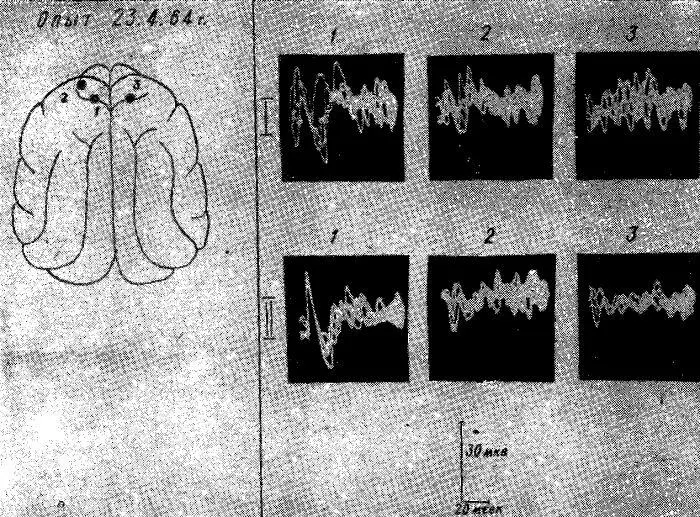
Рис. 14. Изменения характера выявления и распространения соматосензорного вызванного ответа по коре мозга у голодной кошки после внутривенного введения 40% — 2 мл раствора глюкозы. Уретановый наркоз.
I — вызванный ответ в тех же точках 1, 2, 3 до введения глюкозы; II — вызванный ответ в тех же точках через 15 минут после введения глюкозы.
Все это указывало на то, что изменения соматосензорного вызванного ответа у голодного животного определялись восходящими активирующими влияниями гипоталамуса на кору мозга.
В результате этих пищевых возбуждений происходит своеобразное ««вычитание» синапсов из целостной деятельности, участвующей в формировании соматосензорных вызванных потенциалов.
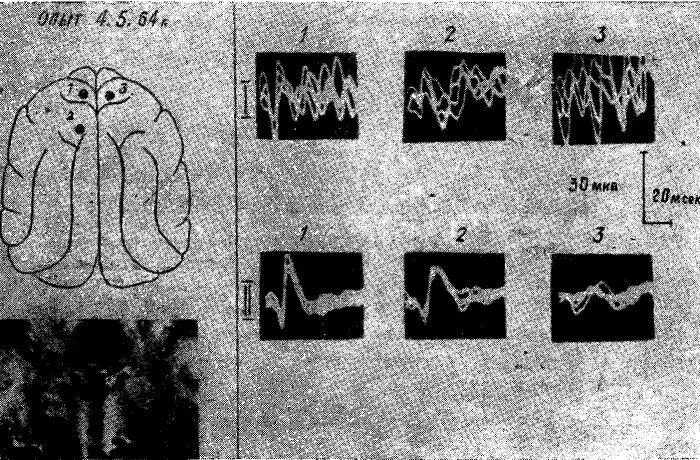
Рис. 15. Изменение характера и распространения соматосензорного вызванного ответа по коре мозга у голодной кошки после коагуляции вентромедиальных отделов гипоталамуса и субталамуса. Уретановый наркоз.
I — исходный вызванный потенциал в точках 1, 2, 3. II — вызванный потенциал в тех же точках после коагуляции медиальных отделов гипоталамуса и субталамуса.
Поскольку в состоянии голода у животных наблюдалось избирательное подавление отрицательной фазы первичного ответа и вторичного положительного колебания соматосензорного вызванного ответа, то можно думать, что возбуждения гипоталамических отделов пищевого центра в состоянии голода адресуются именно к тем синаптическим организациям сензомоторной области коры, которые формируют эти фазы вызванного потенциала. Так как отрицательная фаза первичного ответа, а также вторичные потенциалы обусловливаются восходящими влияниями неспецифических подкорковых систем, адресующихся к аксодендритньим синапсам поверхностных слоев коры (2, 3, 4 и др.), можно думать, что восходящие активирующие влияния гипоталамических отделов пищевого центра специфически адресуются именно к этим синаптическим образованиям коры головного мозга.
Специальные опыты показали, что блокада распространения самотосензорных потенциалов по коре мозга у голодных животных происходит и на уровне медиальных ядер таламуса. Коагуляция медиальных ядер таламуса у накормленных животных прекращает генерализованное распространение вызванных потенциалов на передние отделы контралатерального полушария.
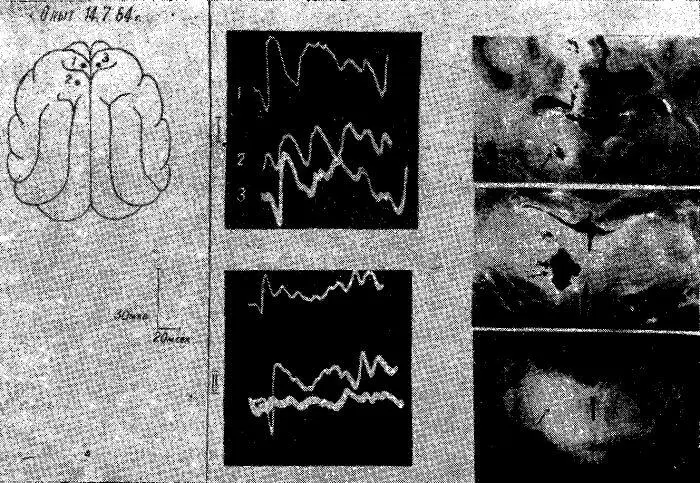
Рис. 16. Изменения распространения соматосензорного вызванного потенциала по коре мозга у накормленной кошки. Уретановый наркоз.
I — до, II — после коагуляции медиальных групп ядер правого таламуса. После этого вызванный потенциал перестает распространяться на ипсилатеральной стороне.
На основании этих опытов мы пришли к выводу, что восходящие активирующие влияния гипоталамуса в состоянии голода специфически распространяются и на аксодендритные синапсы передних отделов коры за счет первичного возбуждения медиальной группы таламических ядер.
Нейрохимические механизмы восходящих активирующих влияний на кору мозга у голодных животных
Наличие у голодных животных системы пищевого возбуждения избирательно объединяющей корковые и подкорковые элементы, поставило перед нами вопрос, какие механизмы производят функциональное объединение различных нервных элементов в единую систему пищевого возбуждения, не блокируемую даже уретаном? Мы предположили, что такое функциональное объединение при пищевом возбуждении может быть основано на едином химическом механизме системы восходящих активирующих влияний гипоталамических пищевых центров на кору головного мозга.
Читать дальшеИнтервал:
Закладка:








