Чарльз Эллис - Эпигенетика

- Название:Эпигенетика
- Автор:
- Жанр:
- Издательство:Техносфера
- Год:2010
- Город:Москва
- ISBN:978-5-94836-257-1
- Рейтинг:
- Избранное:Добавить в избранное
-
Отзывы:
-
Ваша оценка:
Чарльз Эллис - Эпигенетика краткое содержание
Книга ярко и наглядно повествует о новой науке общебиологического значения — эпигенетике, а также об ее отдельных областях. В издании представлено описание разных эпигенетических сигналов и механизмов их реализации, а также собственно феномен, история и концепции эпигенетики, ее отдельные механизмы и пути реализации эпигенетических сигналов в клетке. Авторы различных глав данной книги — ведущие в мире специалисты в области эпигенетики, являющиеся, как правило, и основоположниками ее отдельных областей.
Издание будет полезно широкому кругу читателей, интересующихся коренными проблемами живого мира, сущности жизни и молекулярных механизмов ее проявления.
По формирующейся традиции современной российской научной литературы, оригинальное русскоязычное печатное издание неопрятно переведено, отвратительно вычитано и содержит большое количество ошибок, начиная с обложки. Чарльз Дэвид Эллис указан как С. Д. Эллис.
Эпигенетика - читать онлайн бесплатно полную версию (весь текст целиком)
Интервал:
Закладка:
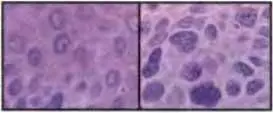
Рис. 24.3.Структурные изменения хроматина в раковых клетках
Эти две микрофотографии сделаны у пациента со сквамозноклеточной карциномой кожи. На левой части показаны нормальные эпидермальные клетки, расположенные в одном миллиметре от прилегающей опухоли и показанные при том же увеличении на правой части рисунка Хроматин, окрашенный пурпурным цветом, благодаря его сродству к гематоксилину, оказывается гораздо более грубым и гранулярным в раковых клетках, чем в нормальном эпидермисе. Такие различия в окраске хроматина используются патологами в качестве диа гностических критериев раковых заболеваний
Таким образом, в настоящее время признается, что критическую роль в канцерогенезе человека играет метилирование цитозина в ДНК. Почти все гены человека содержат остатки метилированного цитозина в своих кодирующих областях, для которых в течение некоторого времени известно непропорциональное участие в образовании болезнетворных мутаций. Метилирование 5 углерода в цитозиновом кольце повышает частоту гидролитического дезаминирования этого основания в двунитевой ДНК. Однако продуктом дезаминирования 5-метилцитозина является в тимин, а не урацил (см. рис. 3.12). Механизмы репарации ДНК впоследствии менее эффективны при восстановлении неправильных сочетаний оснований в ДНК, вызванных дезаминированием. Известно, что метилированные сайты CpG производят более 1/3 всех мутаций типа транзиций, возникающих в зародышевой линии человека (Rideout et al., 1990). Это также справедливо для генов, вызывающих рак, таких как р53 (Rideout et al., 1990). Более удивительным является наблюдение, что этот механизм также принимает существенное участие в образовании инактивирующих мутаций в генах-супрессорах опухоли в соматических тканях. Например, более 50% всех мутаций р53, которые приобретаются при спорадических раковых заболеваниях толстой и прямой кишки, имеют место в сайтах метилирования цитозина (Greenblatt et al., 1994). Таким образом, модификация ДНК ДНК-метилтрансферазами (DNMTs) существенно повышает риск получения рака с помощью такого эндогенного механизма.
Было также показано, что метилирование цитозиновых остатков поддерживает образование канцерогенных аддуктов присоединения между ДНК и канцерогенами, такими как бензопирен из сигаретного дыма. В этом случае, метилирование цитозинового остатка повышает образование канцерогенных аддуктов между соседним гуаниновым остатком и бензопирендиолэпоксидом, приводящее к повышению вероятности мутаций в сайтах CpG в легких курильщиков (Greenblatt et al., 1994; Pfeifer et al., 2000). Интересно, что метилирование может также изменять частоту мутаций в гене р53 в коже, экспонированной солнечному свету (Greenblatt et al., 1994; Pfeifer et al., 2000). Это происходит из-за того, что метиловая группа смещает спектр поглощения цитозина в диапазон падающего солнечного света, тем самым повышая образование пиримидиновых димеров в ДНК клеток кожи. Таким образом, эта эпигенетическая модификация ДНК не только повышает спонтанный мутагенез, но также может влиять на то, как ДНК взаимодействует с канцерогенами и ультрафиолетом (Pfeifer et al., 2000).
Гипометилирование ДНК, которое было давно отмечено в опухолях животных и человека (табл. 24.2), влияет на стабильность хромосом и повышает шанс анеуплоидии. Геномная нестабильность — это признак ракового заболевания, и повышенная ломкость хромосом, вызываемая гипометилированием саттелитных и других последовательностей, может, по-видимому, приводить к образованию раковых заболеваний путем уменьшения стабильности генома (Narayan et al., 1998; Gaudet et al., 2003). Конкретные механизмы, с помощью которых достигается эта нестабильность, все еще не полностью поняты, но они легко могут быть результатом измененных взаимодействий ДНК и белков, вызванных гипометилированием.
4. Гиперметилированные промоторы генов при раковых заболеваниях
4.1. Гены, участвующие в процессе
Наиболее понятный механизм, при помощи которого метилирование ДНК приводит к раковым заболеваниям, это локальное гиперметилирование промоторов генов-супрессоров опухоли. Конкретные механизмы, при помощи которых осуществляется это гиперметилирование, детально обсуждаются ниже. Однако ясно, что это важнейший путь, приводящий к наследуемому сайленсингу генов, которые подавляют развитие рака (Jones and Laird, 1999; Jones and Baylin, 2002; Herman and Baylin, 2003). Обычно, гиперметилирование ДНК осуществляется в участках, обогащенных CpG, или в или CpG-островках, которые локализованы внутри и вокруг транскрипционного стартового сайта аномально «молчащих» генов при раковых заболеваниях. Важно знать, что метилирование цитозина в CpG-островках поблизости от местоположения стартового сайта гена особенно критично, так как эта же самая модификация ДНК, имеющая место внутри генов, обычно не коррелирует с транскрипционным статусом (Jones, 1999).
Перечень связанных с раком генов, на которые влияет упомянутое выше нарушение транскрипции, неуклонно растет. Как было описано ранее, сюда включаются гены в любых положениях на хромосомах (Jones and Baylin, 2002). Действительно, данное эпигенетическое изменение может сейчас численно превосходить те гены, которые часто мутируют в опухолях человека. Как было упомянуто ранее (табл. 24.1), потеря функции гена-супрессора опухоли как результат вызывающего сайленсинг данного гена метилирования CpG, по существу, затрагивает все известные пути. Чтобы понять значение генов для процесса канцерогенеза и сформулировать проблему в этой области на будущее, гены, по-видимому, могут быть разделены на три группы.
Первая группа содержит те гены, которые участвуют в определении гиперметилирования промотора и сайленсинга генов, как одного из важных механизмов потери функции гена-супрессора опухоли при раковых заболеваниях (табл. 24.3). Эти гены уже были распознаны как классические гены-супрессоры опухоли, которые при мутации в зародышевой линии семьи вызывают наследуемые формы рака (Jones and Laird, 1999; Jones and Baylin, 2002; Herman and Baylin, 2003). Они также часто мутируют при спорадических формах рака, но в таких опухолях они нередко могут быть гиперметилированы по одной или обеим аллелям (Jones and Laird, 1999; Jones and Baylin, 2002; Herman and Baylin, 2003). Кроме того, для этих генов гиперметилирование промотора может иногда составлять «второй удар» (по гипотезе Кнудсона) в связи с тем, что они ассоциированы с потерей функции второй копии этого гена при семейных опухолях, где «первым ударом» является мутация в зародышевой линии (Grady et al., 2000; Esteller et al., 2001b). В некоторых случаях было показано, что индуцированная 5-азацитидином реактивация этих генов в культуре опухолевых клеток восстанавливает ключевую функцию гена-супрессора опухоли, утраченную в процессе роста опухоли. Таким примером является функция коррекции неправильного спаривания оснований ДНК в клетках рака толстой кишки, где имеет место сайленсинг гена MLH1 (Herman et al., 1998).
Читать дальшеИнтервал:
Закладка:



